

Force generation by a dynamic Z-ring in Escherichia coli cell division
- PMID: 19114664
- PMCID: PMC2629190
- DOI: 10.1073/pnas.0808657106
Force generation by a dynamic Z-ring in Escherichia coli cell division
Abstract
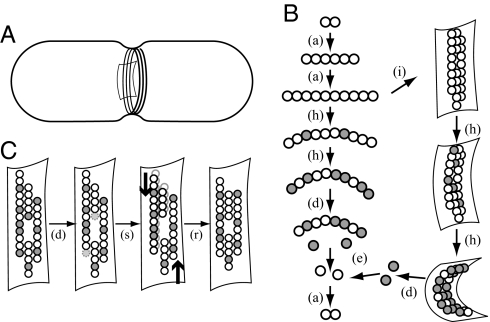
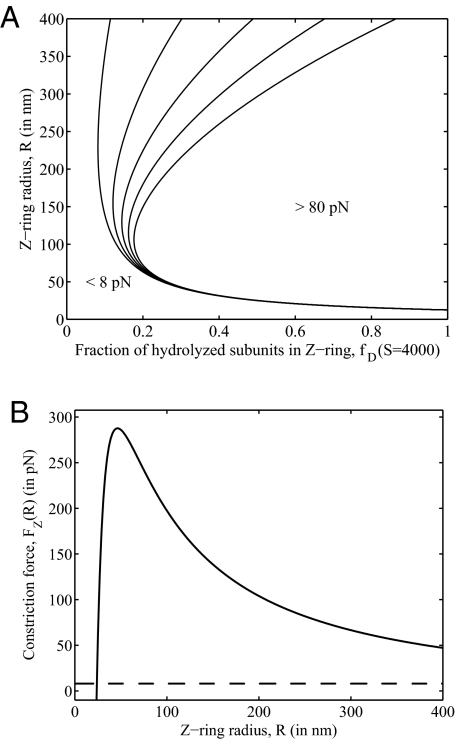
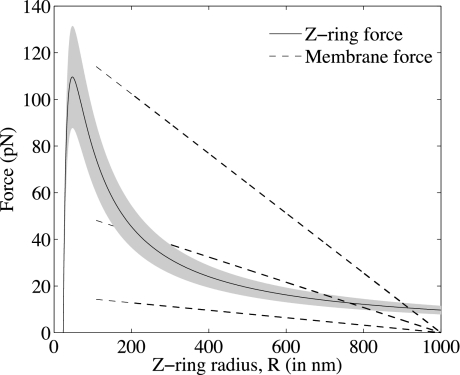
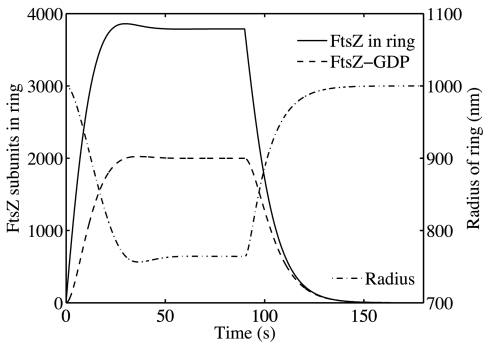
FtsZ, a bacterial homologue of tubulin, plays a central role in bacterial cell division. It is the first of many proteins recruited to the division site to form the Z-ring, a dynamic structure that recycles on the time scale of seconds and is required for division to proceed. FtsZ has been recently shown to form rings inside tubular liposomes and to constrict the liposome membrane without the presence of other proteins, particularly molecular motors that appear to be absent from the bacterial proteome. Here, we propose a mathematical model for the dynamic turnover of the Z-ring and for its ability to generate a constriction force. Force generation is assumed to derive from GTP hydrolysis, which is known to induce curvature in FtsZ filaments. We find that this transition to a curved state is capable of generating a sufficient force to drive cell-wall invagination in vivo and can also explain the constriction seen in the in vitro liposome experiments. Our observations resolve the question of how FtsZ might accomplish cell division despite the highly dynamic nature of the Z-ring and the lack of molecular motors.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Vaughan S, Wickstead B, Gull K, Addinall S. Molecular evolution of FtsZ protein sequences encoded within the genomes of archaea, bacteria, and eukaryota. J Mol Evol. 2004;58:19–29. - PubMed
-
- Miyagishima S. Origin and evolution of the chloroplast division machinery. J Plant Res. 2005;118:295–306. - PubMed
-
- Bi E, Lutkenhaus J. FtsZ ring structure associated with division in Escherichia coli. Nature. 1991;354:161–164. - PubMed
-
- Lutkenhaus J. FtsZ ring in bacterial cytokinesis. Mol Microbiol. 1993;9:403–409. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
