

A recalibrated molecular clock and independent origins for the cholera pandemic clones
- PMID: 19115014
- PMCID: PMC2605724
- DOI: 10.1371/journal.pone.0004053
A recalibrated molecular clock and independent origins for the cholera pandemic clones
Abstract
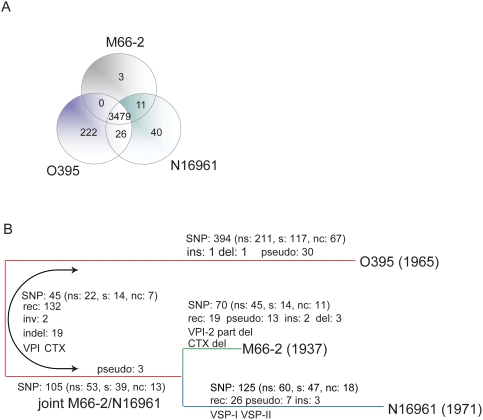
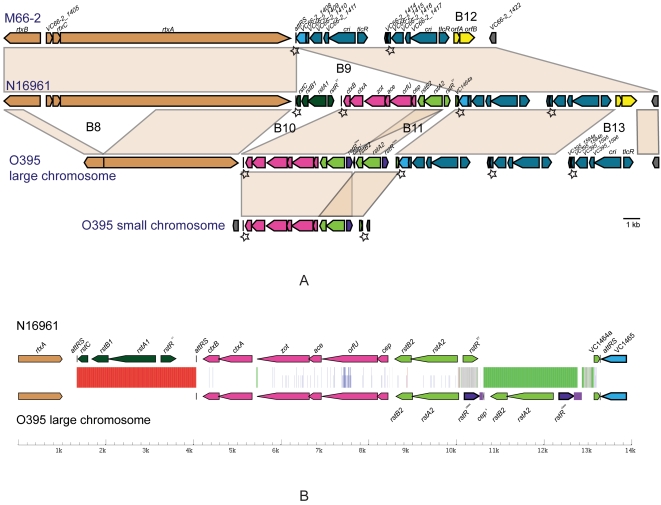
Cholera, caused by Vibrio cholerae, erupted globally from South Asia in 7 pandemics, but there were also local outbreaks between the 6(th) (1899-1923) and 7(th) (1961-present) pandemics. All the above are serotype O1, whereas environmental or invertebrate isolates are antigenically diverse. The pre 7th pandemic isolates mentioned above, and other minor pathogenic clones, are related to the 7(th) pandemic clone, while the 6(th) pandemic clone is in the same lineage but more distantly related, and non-pathogenic isolates show no clonal structure. To understand the origins and relationships of the pandemic clones, we sequenced the genomes of a 1937 prepandemic strain and a 6(th) pandemic isolate, and compared them with the published 7(th) pandemic genome. We distinguished mutational and recombinational events, and allocated these and other events, to specific branches in the evolutionary tree. There were more mutational than recombinational events, but more genes, and 44 times more base pairs, changed by recombination. We used the mutational single-nucleotide polymorphisms and known isolation dates of the prepandemic and 7(th) pandemic isolates to estimate the mutation rate, and found it to be 100 fold higher than usually assumed. We then used this to estimate the divergence date of the 6(th) and 7(th) pandemic clones to be about 1880. While there is a large margin of error, this is far more realistic than the 10,000-50,000 years ago estimated using the usual assumptions. We conclude that the 2 pandemic clones gained pandemic potential independently, and overall there were 29 insertions or deletions of one or more genes. There were also substantial changes in the major integron, attributed to gain of individual cassettes including copying from within, or loss of blocks of cassettes. The approaches used open up new avenues for analysing the origin and history of other important pathogens.
Conflict of interest statement
Figures




References
-
- Blake PA. Historical perspectives on pandemic cholera. In: Wachsmuth IK, Blake PA, Olsvik O, editors. Vibrio cholerae and cholera, molecular to global perspectives. Washington: ASM Press; 1994. pp. 293–295.
-
- Mukerjee S. Problems of Cholera (El Tor). Am J Trop Med Hyg. 1963;12:388–392. - PubMed
-
- Reeves PR, Lan R. Cholera in the 1990s. Br Med Bull. 1998;54:611–623. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
