

Characterization of relaxin receptor (RXFP1) desensitization and internalization in primary human decidual cells and RXFP1-transfected HEK293 cells
- PMID: 19116340
- PMCID: PMC2671891
- DOI: 10.1210/en.2008-1385
Characterization of relaxin receptor (RXFP1) desensitization and internalization in primary human decidual cells and RXFP1-transfected HEK293 cells
Abstract
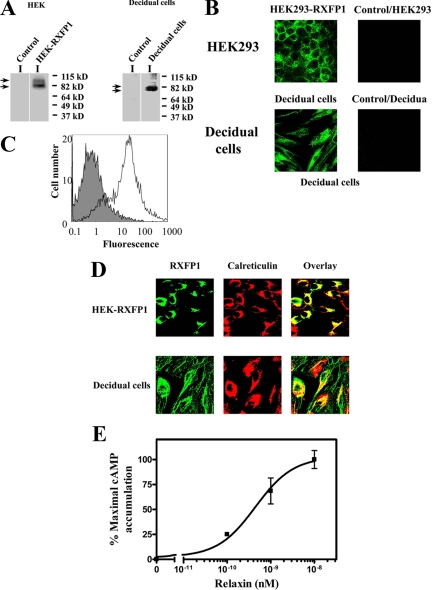
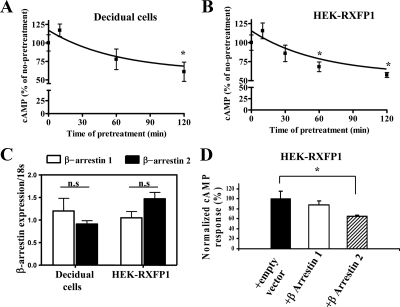
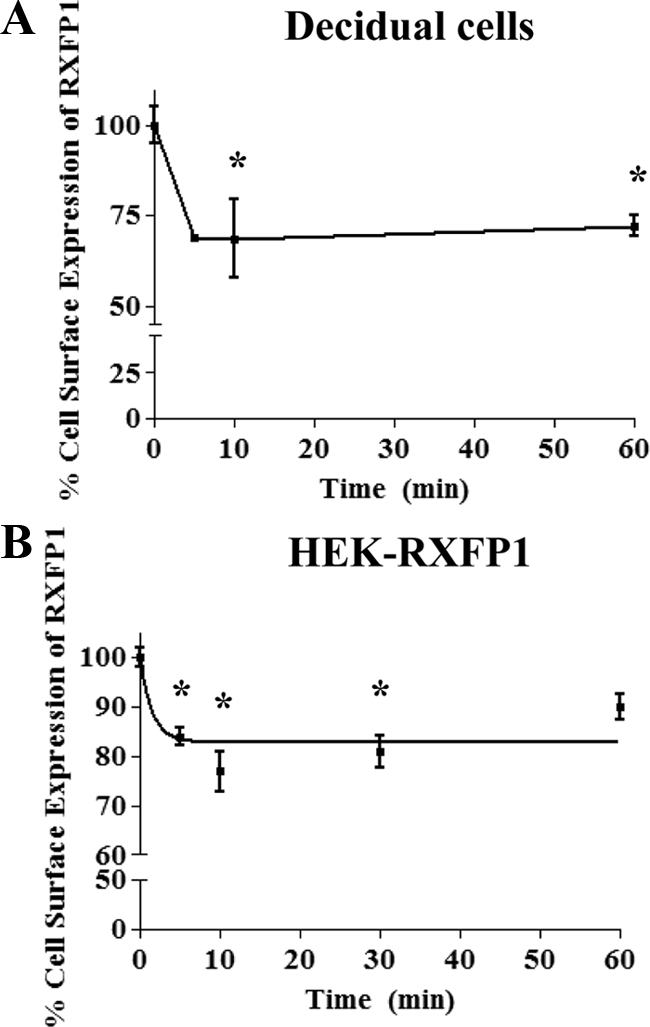
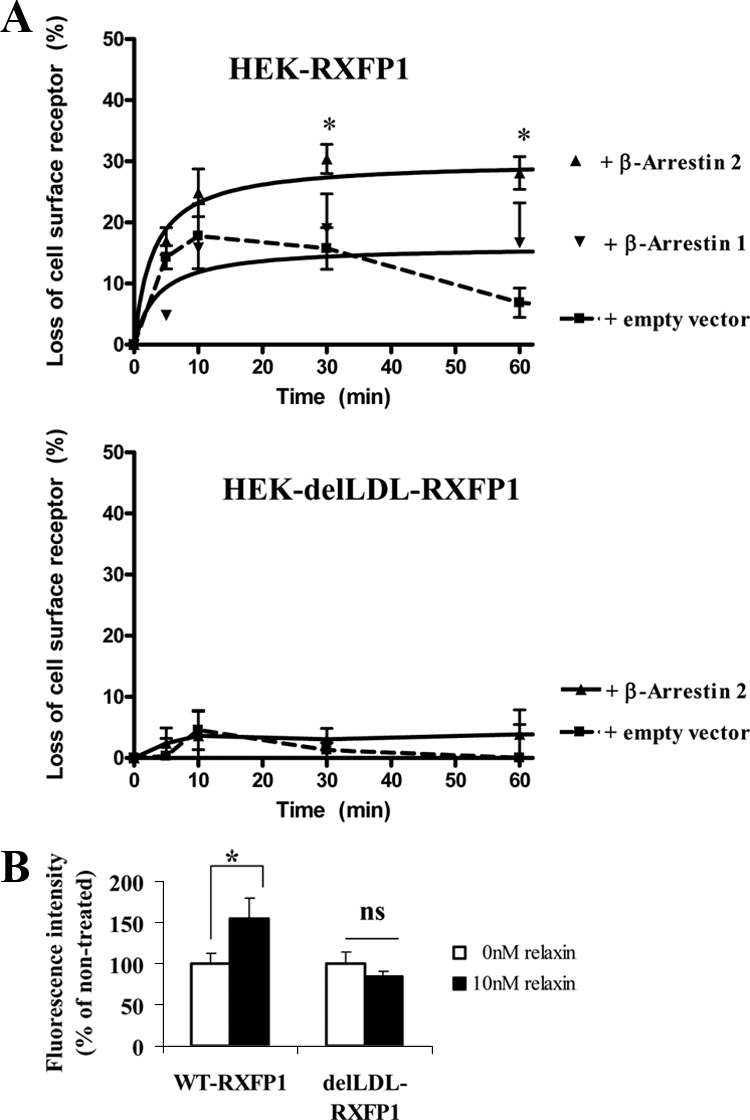
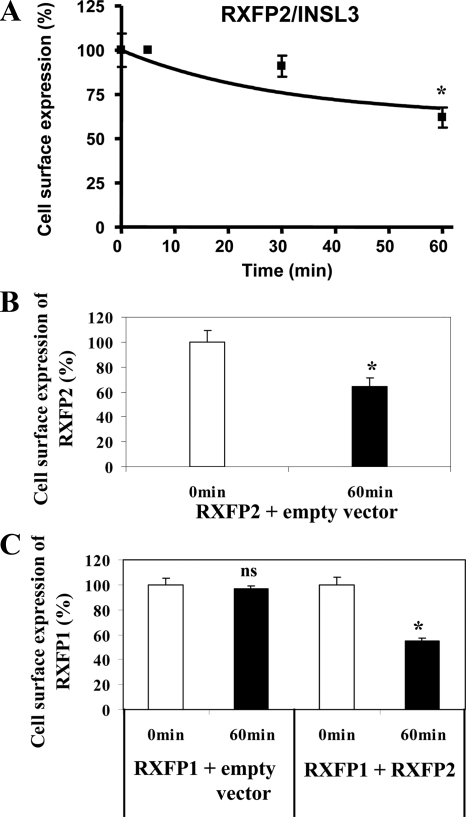
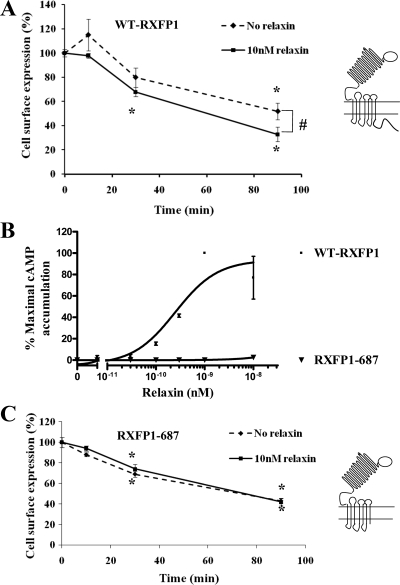
We report here the desensitization and internalization of the relaxin receptor (RXFP1) after agonist activation in both primary human decidual cells and HEK293 cells stably transfected with RXFP1. The importance of beta-arrestin 2 in these processes has also been demonstrated. Thus, in HEK-RXFP1 cells the desensitization of RXFP1 was significantly increased when beta-arrestin 2 was overexpressed. After relaxin activation, beta-arrestin 2 was translocated to the cell membrane and RXFP1 underwent rapid internalization. We have previously shown that RXFP1 forms dimers/oligomers during its biosynthesis and trafficking to the plasma membrane, we now show that internalization of RXFP1 occurs through this dimerization/oligomerization. In nonagonist stimulated cells, it is known that the majority of the RXFP1 is located intracellularly and was confirmed in the cells used here. Constitutive internalization of RXFP1 could account for this and indeed, slow but robust constitutive internalization, which was increased after agonist stimulation was demonstrated. A carboxyl-terminal deleted RXFP1 variant had a similar level of constitutive agonist-independent internalization as the wild-type RXFP1 but lost sensitivity to agonist stimulation. This demonstrated the importance of the carboxyl terminus in agonist-stimulated receptor internalization. These data suggest that the autocrine/paracrine actions of relaxin in the decidua are under additional controls at the level of expression of its receptor on the surface of its target cells.
Figures
References
-
- James R, Niall H, Kwok S, Bryant-Greenwood GD 1977 Primary structure of porcine relaxin: homology with insulin and related growth factors. Nature 267:544–546 - PubMed
-
- Sherwood OD 2004 Relaxin’s physiological roles and other diverse actions. Endocr Rev 25:205–234 - PubMed
-
- Samuel CS, Hewitson TD 2006 Relaxin in cardiovascular and renal disease. Kidney Int 69:1498–1502 - PubMed
-
- Mookerjee I, Solly NR, Royce SG, Tregear GW, Samuel CS, ML. T 2006 Endogenous relaxin regulates collagen deposition in an animal model of allergic airway disease. Endocrinology 147:754–761 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
