

Dystrophin glycoprotein complex-associated Gbetagamma subunits activate phosphatidylinositol-3-kinase/Akt signaling in skeletal muscle in a laminin-dependent manner
- PMID: 19117013
- PMCID: PMC2820875
- DOI: 10.1002/jcp.21684
Dystrophin glycoprotein complex-associated Gbetagamma subunits activate phosphatidylinositol-3-kinase/Akt signaling in skeletal muscle in a laminin-dependent manner
Abstract
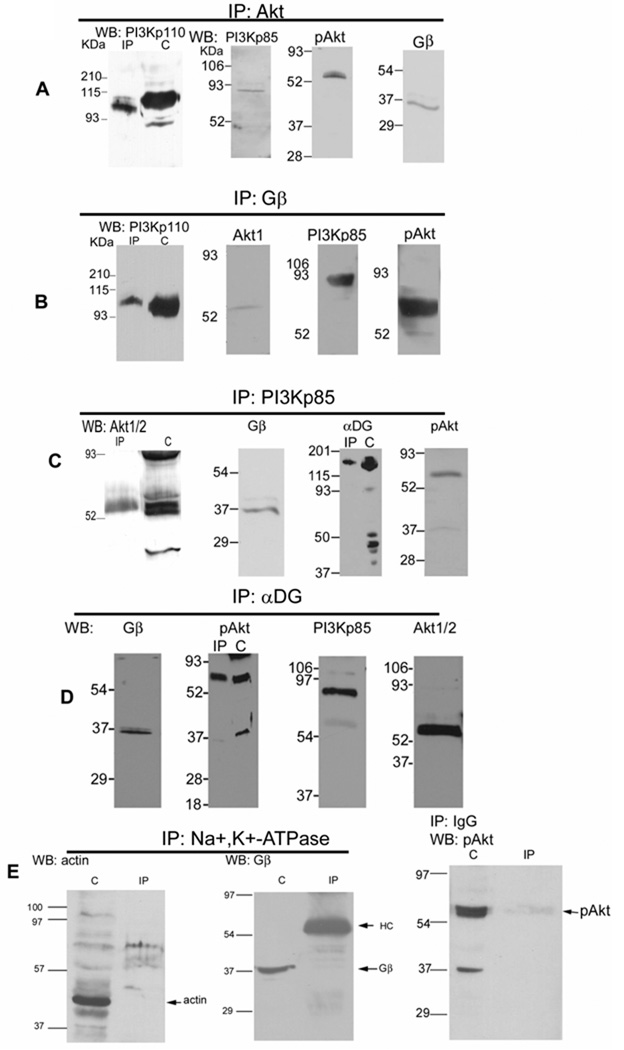
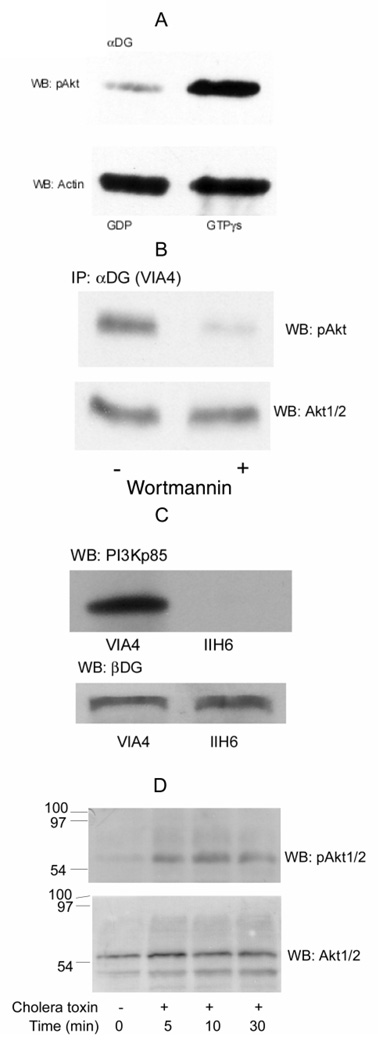
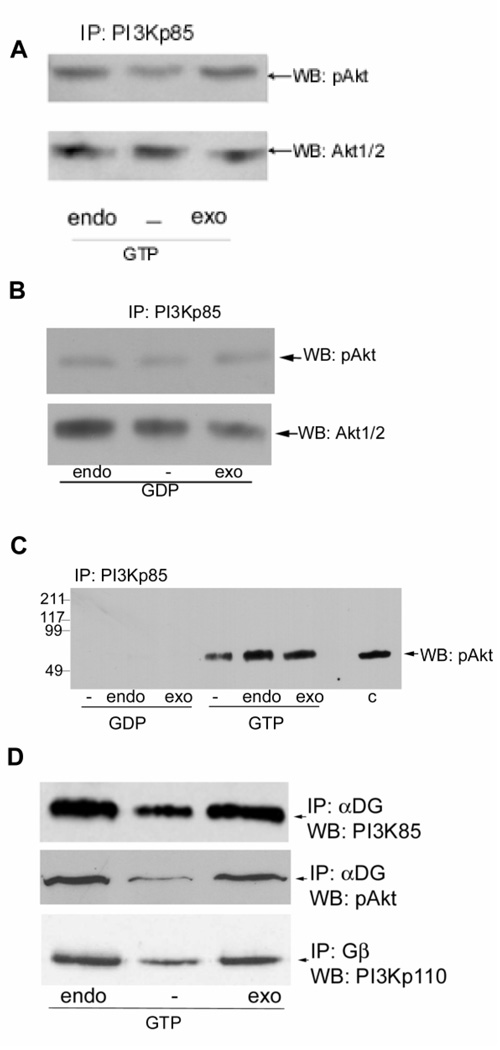
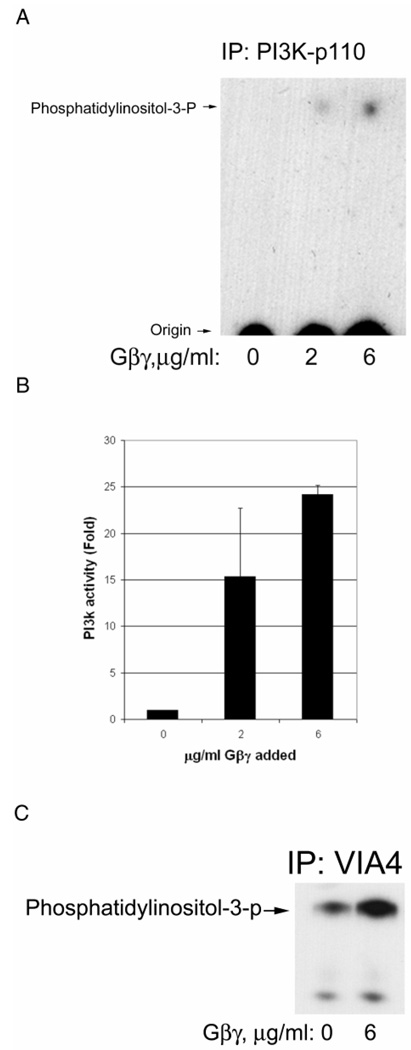
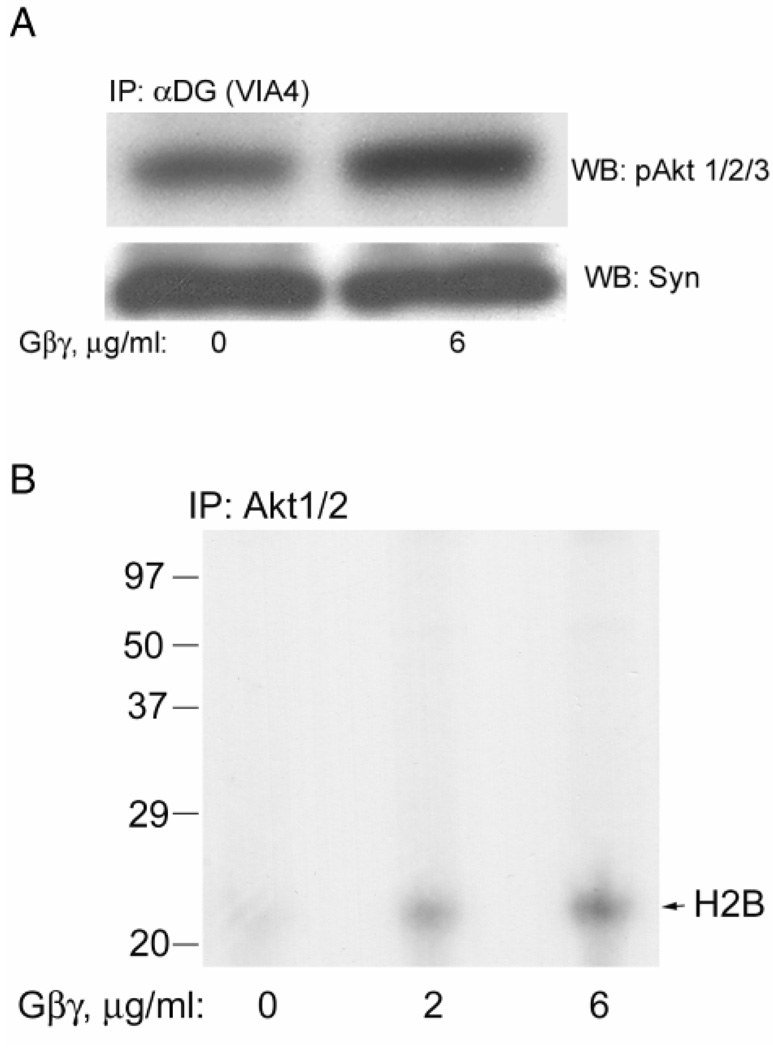
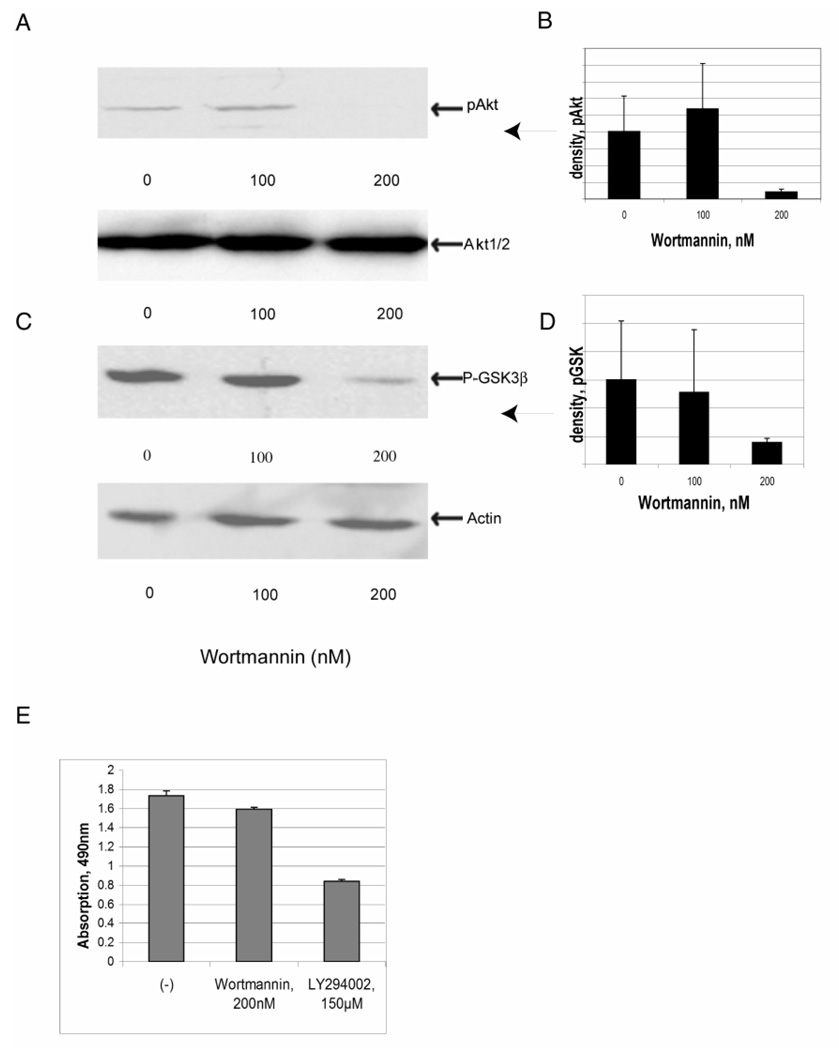
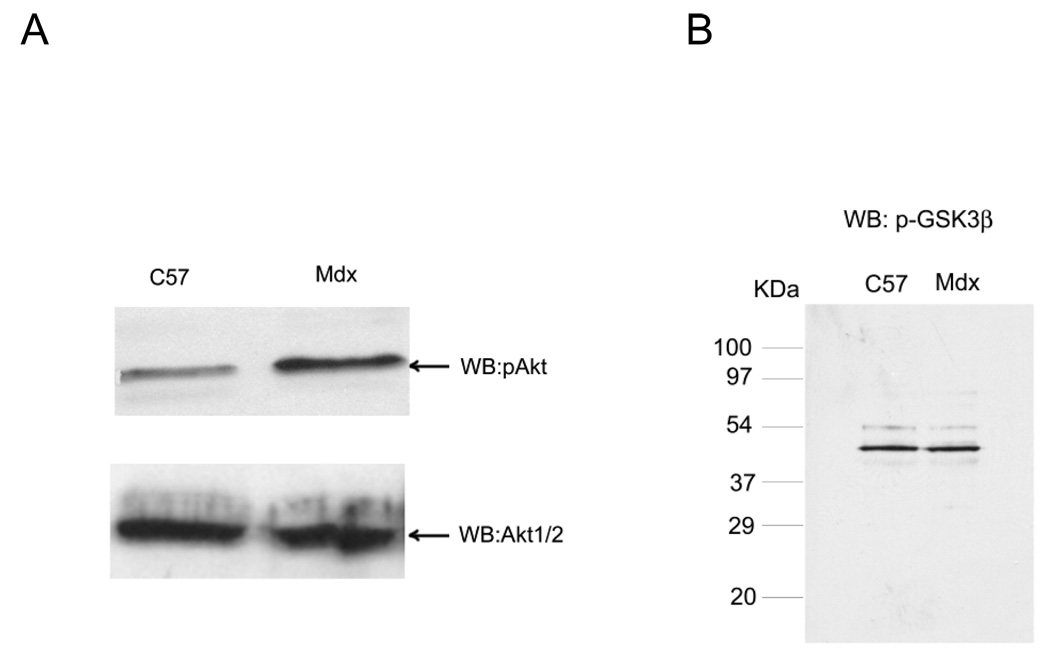
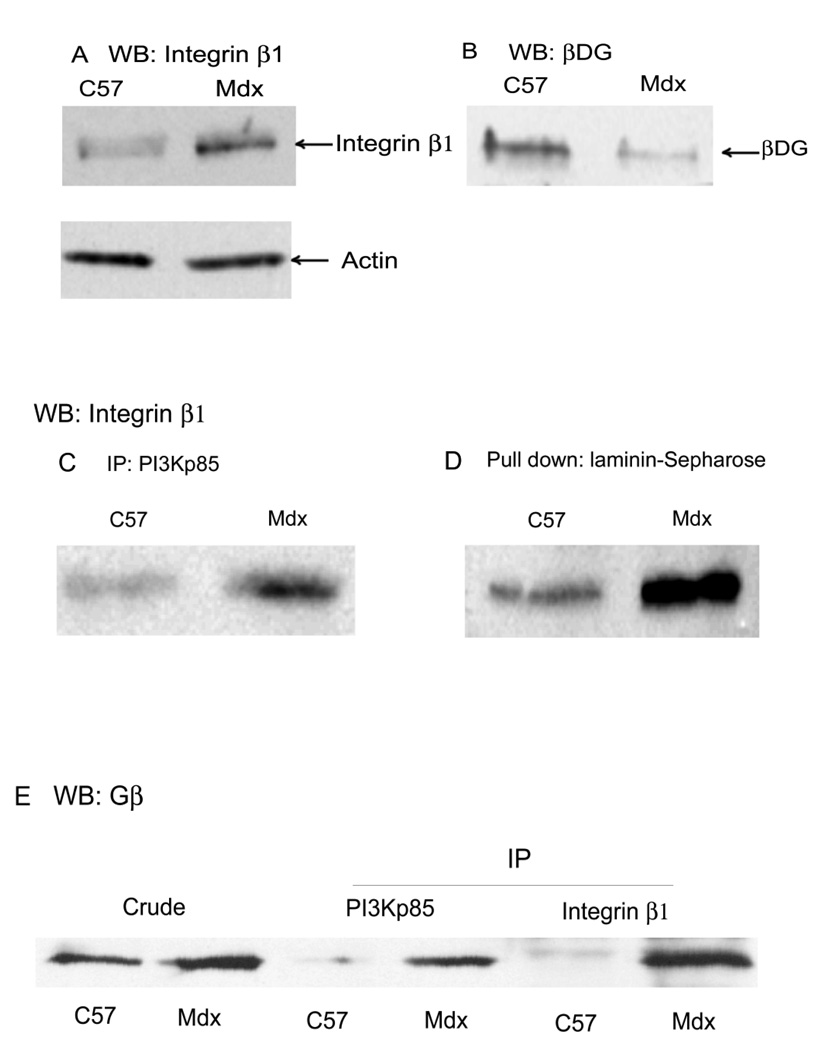
Previously, we showed that laminin-binding to the dystrophin glycoprotein complex (DGC) of skeletal muscle causes a heterotrimeric G-protein (Galphabetagamma) to bind, changing the activation state of the Gsalpha subunit. Others have shown that laminin-binding to the DGC also leads to Akt activation. Gbetagamma, released when Gsalpha is activated, is known to bind phosphatidylinositol-3-kinase (PI3K), which activates Akt in other cells. Here, we investigate whether muscle Akt activation results from Gbetagamma, using immunoprecipitation and immunoblotting, and purified Gbetagamma. In the presence of laminin, PI3K-binding to the DGC increases and Akt becomes phosphorylated and activated (pAkt), and glycogen synthase kinase is phosphorylated. Antibodies, which specifically block laminin-binding to alpha-dystroglycan, prevent PI3K-binding to the DGC. Purified bovine brain Gbetagamma also caused PI3K and Akt activation. These results show that DGC-Gbetagamma is binding PI3K and activating pAkt in a laminin-dependent manner. Mdx mice, which have greatly diminished amounts of DGC proteins, display elevated pAkt signaling and increased expression of integrin beta1 compared to normal muscle. This integrin binds laminin, Gbetagamma, and PI3K. Collectively, these suggest that PI3K is an important target for the Gbetagamma, which normally binds to DGC syntrophin, and activates PI3K/Akt signaling. Disruption of the DGC in mdx mouse is causing dis-regulation of the laminin-DGC-Gbetagamma-PI3K-Akt signaling and is likely to be important to the pathogenesis of muscular dystrophy. Upregulating integrin beta1 expression and activating the PI3K/Akt pathway in muscular dystrophy may partially compensate for the loss of the DGC. The results suggest new therapeutic approaches to muscle disease.
Figures
Similar articles
-
The dystrophin-glycoprotein complex in the prevention of muscle damage.J Biomed Biotechnol. 2011;2011:210797. doi: 10.1155/2011/210797. Epub 2011 Oct 5. J Biomed Biotechnol. 2011. PMID: 22007139 Free PMC article. Review.
-
Laminin-alpha1 globular domains 3 and 4 induce heterotrimeric G protein binding to alpha-syntrophin's PDZ domain and alter intracellular Ca2+ in muscle.Am J Physiol Cell Physiol. 2005 Feb;288(2):C377-88. doi: 10.1152/ajpcell.00279.2004. Epub 2004 Sep 22. Am J Physiol Cell Physiol. 2005. PMID: 15385269
-
Laminin-α1 LG4-5 domain binding to dystroglycan mediates muscle cell survival, growth, and the AP-1 and NF-κB transcription factors but also has adverse effects.Am J Physiol Cell Physiol. 2012 Mar 15;302(6):C902-14. doi: 10.1152/ajpcell.00118.2011. Epub 2011 Dec 7. Am J Physiol Cell Physiol. 2012. PMID: 22159078 Free PMC article.
-
Inhibition of dystroglycan binding to laminin disrupts the PI3K/AKT pathway and survival signaling in muscle cells.Muscle Nerve. 2002 Nov;26(5):644-53. doi: 10.1002/mus.10258. Muscle Nerve. 2002. PMID: 12402286
-
Taking the heart failure battle inside the cell: small molecule targeting of Gβγ subunits.J Mol Cell Cardiol. 2011 Oct;51(4):462-7. doi: 10.1016/j.yjmcc.2011.01.006. Epub 2011 Jan 21. J Mol Cell Cardiol. 2011. PMID: 21256851 Free PMC article. Review.
Cited by
-
Dg-Dys-Syn1 signaling in Drosophila regulates the microRNA profile.BMC Cell Biol. 2012 Oct 29;13:26. doi: 10.1186/1471-2121-13-26. BMC Cell Biol. 2012. PMID: 23107381 Free PMC article.
-
Stress-induced ECM alteration modulates cellular microRNAs that feedback to readjust the extracellular environment and cell behavior.Front Genet. 2013 Dec 31;4:305. doi: 10.3389/fgene.2013.00305. Front Genet. 2013. PMID: 24427166 Free PMC article. Review.
-
Immobilization and therapeutic passive stretching generate thickening and increase the expression of laminin and dystrophin in skeletal muscle.Braz J Med Biol Res. 2014 Jun;47(6):483-91. doi: 10.1590/1414-431x20143521. Epub 2014 May 9. Braz J Med Biol Res. 2014. PMID: 24820070 Free PMC article.
-
Spatiotemporal Mapping Reveals Regional Gastrointestinal Dysfunction in mdx Dystrophic Mice Ameliorated by Oral L-arginine Supplementation.J Neurogastroenterol Motil. 2020 Jan 30;26(1):133-146. doi: 10.5056/jnm19029. J Neurogastroenterol Motil. 2020. PMID: 31715094 Free PMC article.
-
The dystrophin-glycoprotein complex in the prevention of muscle damage.J Biomed Biotechnol. 2011;2011:210797. doi: 10.1155/2011/210797. Epub 2011 Oct 5. J Biomed Biotechnol. 2011. PMID: 22007139 Free PMC article. Review.
References
-
- Baar K, Esser K. Phosphorylation of p70(S6k) correlates with increased skeletal muscle mass following resistance exercise. Am J Physiol. 1999;276(1 Pt 1):C120–C127. - PubMed
-
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. - PubMed
-
- Brown SC, Fassati A, Popplewell L, Page AM, Henry MD, Campbell KP, Dickson G. Dystrophic phenotype induced in vitro by antibody blockade of muscle alpha-dystroglycan-laminin interaction. J Cell Sci. 1999;112(Pt 2):209–216. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
