

Role of TLS DNA polymerases eta and kappa in processing naturally occurring structured DNA in human cells
- PMID: 19117014
- PMCID: PMC2696892
- DOI: 10.1002/mc.20509
Role of TLS DNA polymerases eta and kappa in processing naturally occurring structured DNA in human cells
Abstract
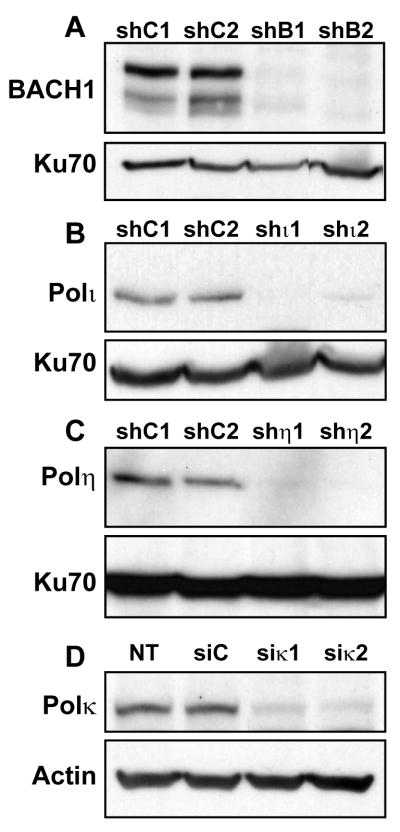
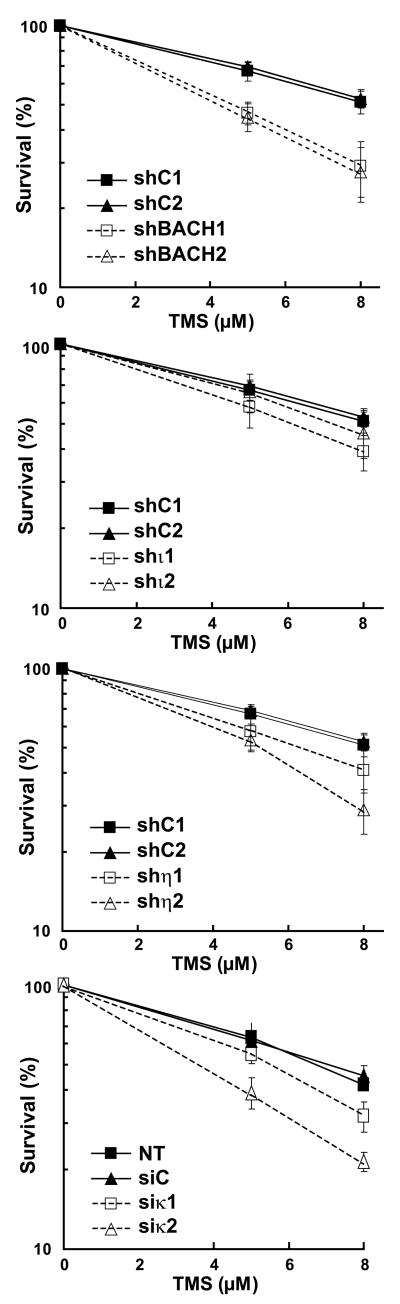
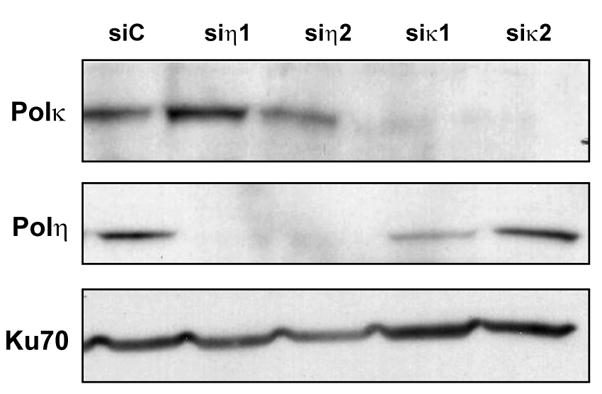
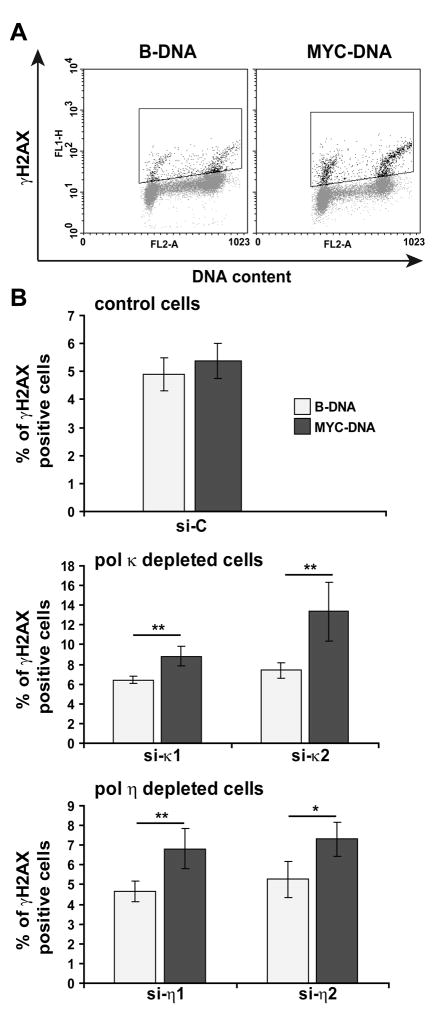
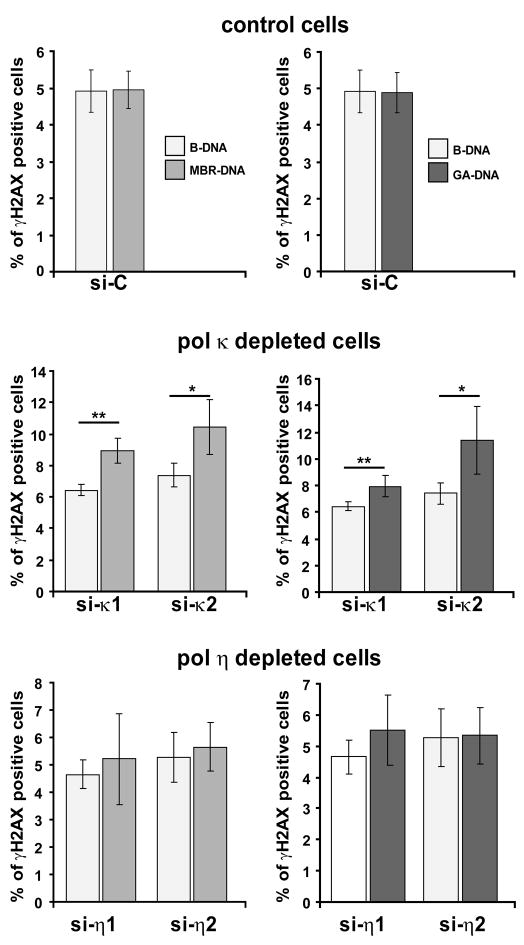
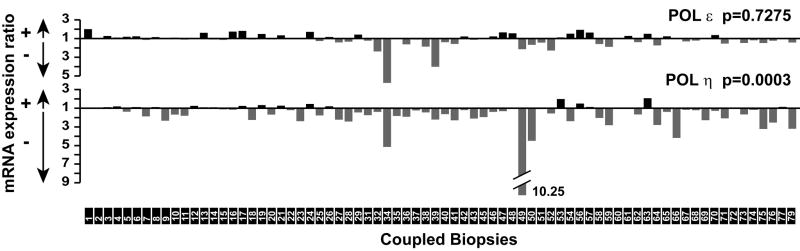
Accurate DNA replication during S-phase is fundamental to maintain genome integrity. During this critical process, replication forks frequently encounter obstacles that impede their progression. While the regulatory pathways which act in response to exogenous replication stress are beginning to emerge, the mechanisms by which fork integrity is maintained at naturally occurring endogenous replication-impeding sequences remains obscure. Notably, little is known about how cells replicate through special chromosomal regions containing structured non-B DNA, for example, G4 quartets, known to hamper fork progression or trigger chromosomal rearrangements. Here, we have investigated the role in this process of the human translesion synthesis (TLS) DNA polymerases of the Y-family (pol eta, pol iota, and pol kappa), specialized enzymes known to synthesize DNA through DNA damage. We show that depletion by RNA interference of expression of the genes for Pol eta or Pol kappa, but not Pol iota, sensitizes U2OS cells treated with the G4-tetraplex interactive compound telomestatin and triggers double-strand breaks in HeLa cells harboring multiple copies of a G-rich sequence from the promoter region of the human c-MYC gene, chromosomally integrated as a transgene. Moreover, we found that downregulation of Pol kappa only raises the level of DSB in HeLa cells containing either one of two breakage hotspot structured DNA sequences in the chromosome, the major break region (Mbr) of BCL-2 gene and the GA rich region from the far right-hand end of the genome of the Kaposi Sarcoma associated Herpesvirus. These data suggest that naturally occurring DNA structures are physiological substrates of both pol eta and pol kappa. We discuss these data in the light of their downregulation in human cancers.
(c) 2008 Wiley-Liss, Inc.
Figures
References
-
- Diffley JF. Regulation of early events in chromosome replication. Curr Biol. 2004;14(18):R778–786. - PubMed
-
- Kolodner RD, Putnam CD, Myung K. Maintenance of genome stability in Saccharomyces cerevisiae. Science. 2002;297(5581):552–557. - PubMed
-
- Burhans WC, Carr AM, Wahl GM. DNA Replication and Cancer. In: DePamphilis ML, editor. DNA Replication and Human Disease. Cold Spring Harbor Laboratory press; 2006. pp. 481–500.
-
- Bacolla A, Wojciechowska M, Kosmider B, Larson JE, Wells RD. The involvement of non-B DNA structures in gross chromosomal rearrangements. DNA Repair (Amst) 2006;5(9–10):1161–1170. - PubMed
-
- Bowater RP, Wells RD. The intrinsically unstable life of DNA triplet repeats associated with human hereditary disorders. Prog Nucleic Acid Res Mol Biol. 2001;66:159–202. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
