

A mathematical model of murine metabolic regulation by leptin: energy balance and defense of a stable body weight
- PMID: 19117546
- PMCID: PMC2643084
- DOI: 10.1016/j.cmet.2008.11.005
A mathematical model of murine metabolic regulation by leptin: energy balance and defense of a stable body weight
Abstract
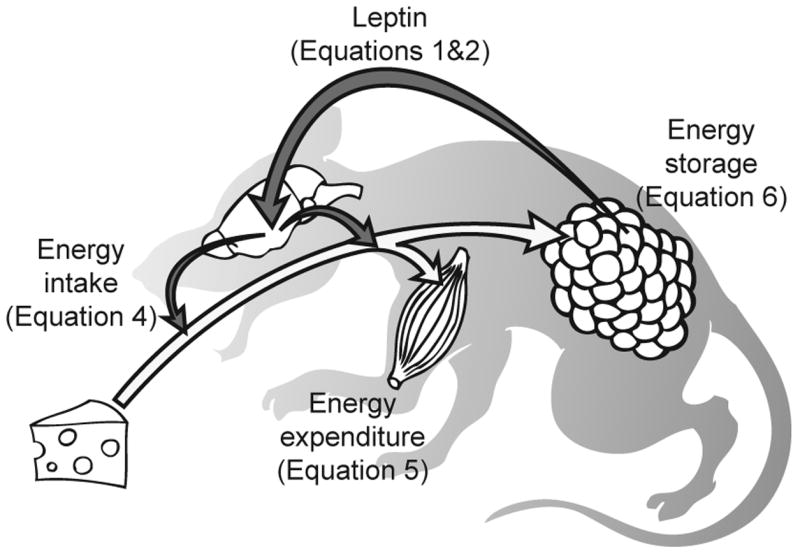
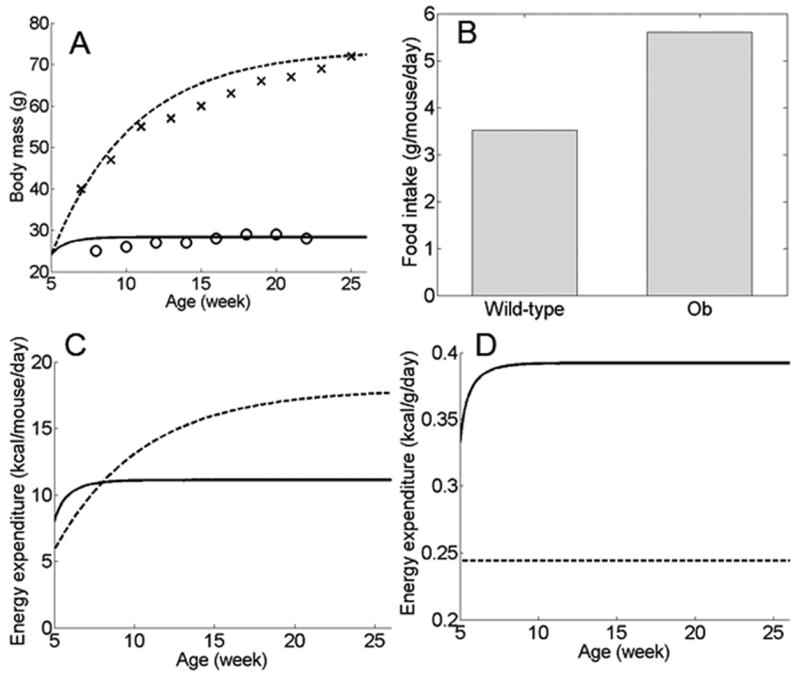
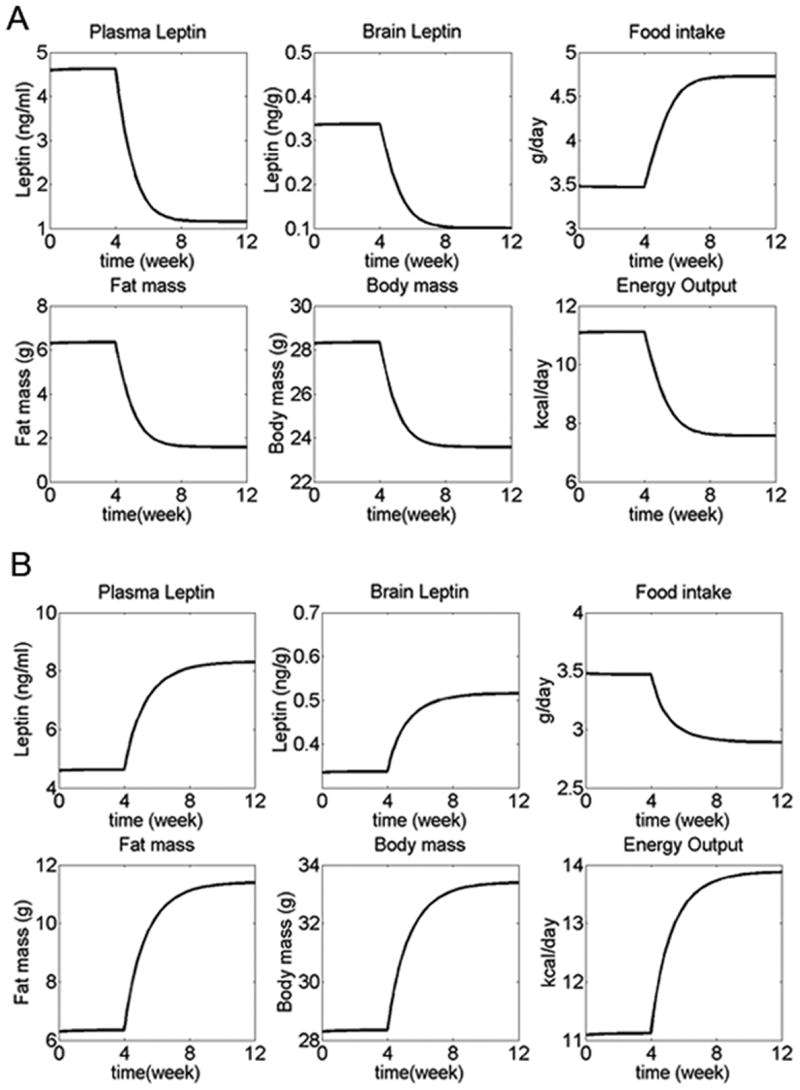
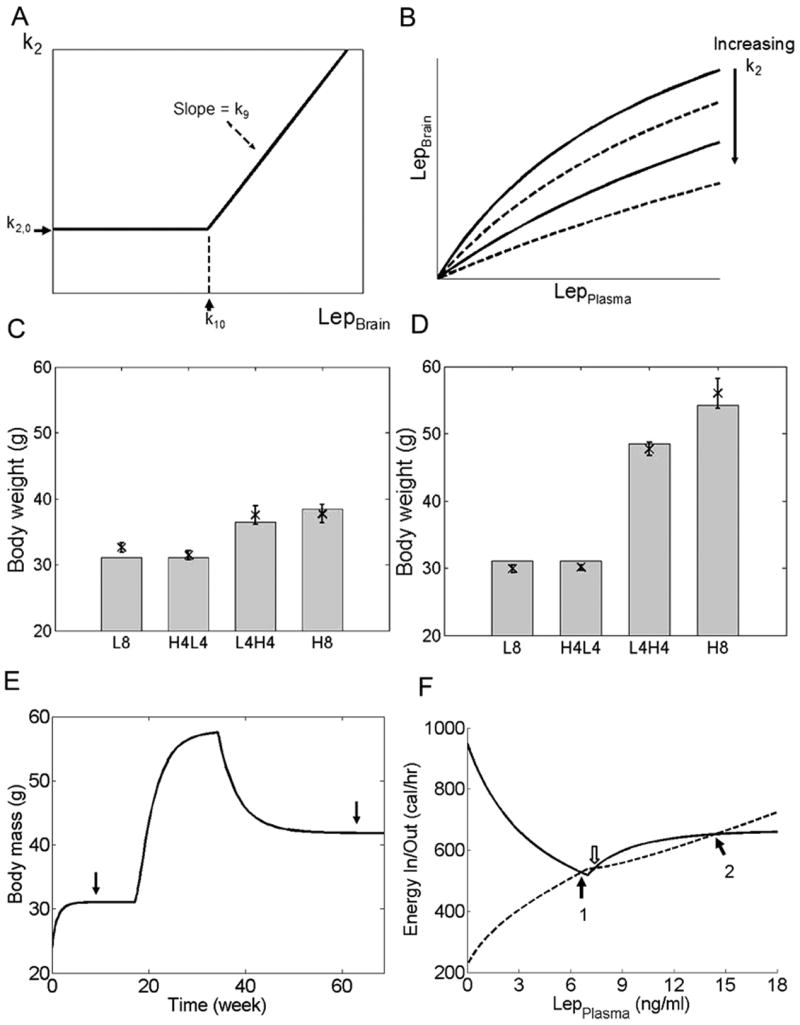
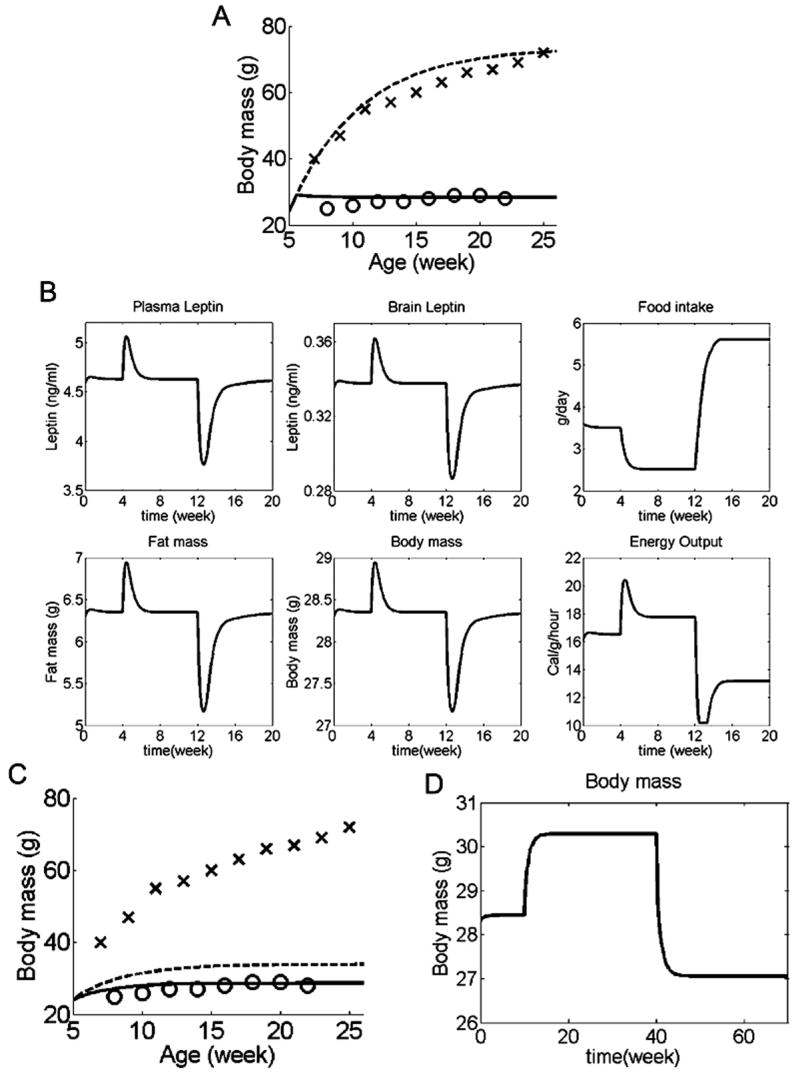
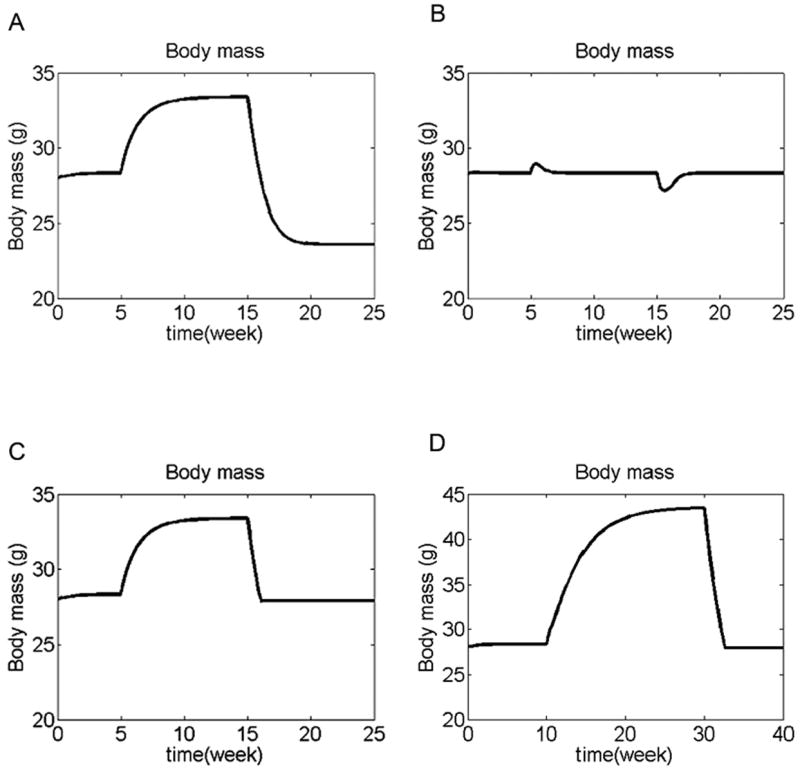
We have developed a physiologically based mathematical model, with parameters derived from published experimental data, to simulate the regulatory effects of the leptin pathway on murine energy homeostasis. Model outcomes are consistent with data reported in the literature and reproduce key characteristics of the energy regulatory system, including compensatory responses that counteract changes in body weight and the failure of this ability when the leptin pathway is disrupted. Our model revealed the possibility of multiple steady states for body weight. It also provided a unified theoretical framework for two historically antagonistic hypotheses regarding body weight regulation ("set-point" versus "settling point"). Finally, our model has identified potential avenues for future investigations.
Figures
Comment in
-
Models use leptin and calculus to count calories.Cell Metab. 2009 Jan 7;9(1):3-4. doi: 10.1016/j.cmet.2008.12.006. Cell Metab. 2009. PMID: 19117539
References
-
- Abdel-Hamid TK. Exercise and diet in obesity treatment: an integrative system dynamics perspective. Med Sci Sports Exerc. 2003;35:400–413. - PubMed
-
- Ahima RS, Prabakaran D, Mantzoros C, Qu D, Lowell B, Maratos-Flier E, Flier JS. Role of leptin in the neuroendocrine response to fasting. Nature. 1996;382:250–252. - PubMed
-
- Badman MK, Flier JS. The adipocyte as an active participant in energy balance and metabolism. Gastroenterology. 2007;132:2103–2115. - PubMed
-
- Banks WA, Clever CM, Farrell CL. Partial saturation and regional variation in the blood-to-brain transport of leptin in normal weight mice. Am J Physiol Endocrinol Metab. 2000;278:E1158–1165. - PubMed
-
- Banks WA, DiPalma CR, Farrell CL. Impaired transport of leptin across the blood-brain barrier in obesity. Peptides. 1999;20:1341–1345. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
