

The nuclear orphan receptor COUP-TFII plays an essential role in adipogenesis, glucose homeostasis, and energy metabolism
- PMID: 19117548
- PMCID: PMC2630393
- DOI: 10.1016/j.cmet.2008.12.002
The nuclear orphan receptor COUP-TFII plays an essential role in adipogenesis, glucose homeostasis, and energy metabolism
Abstract
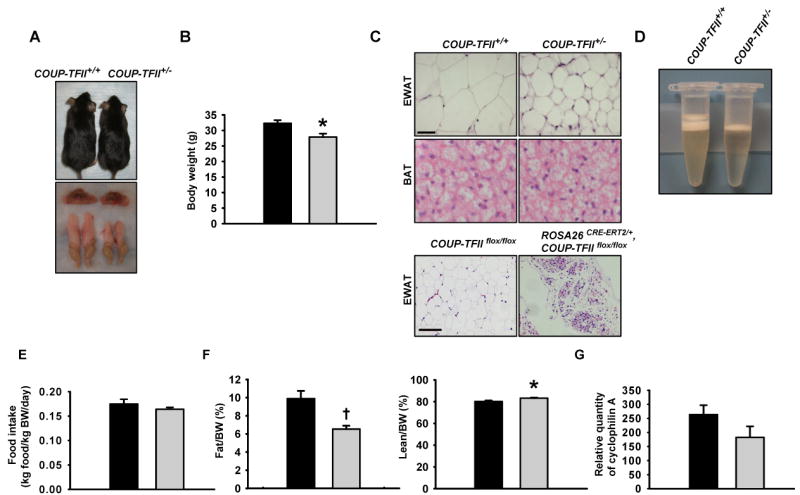
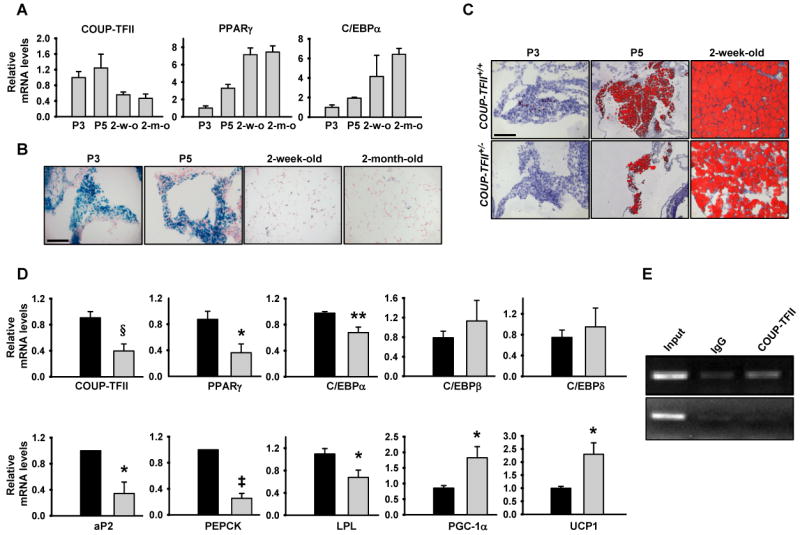
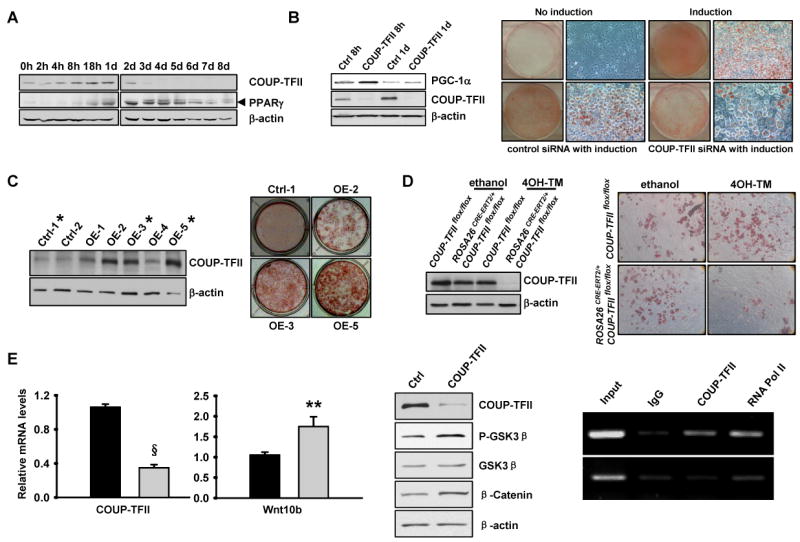
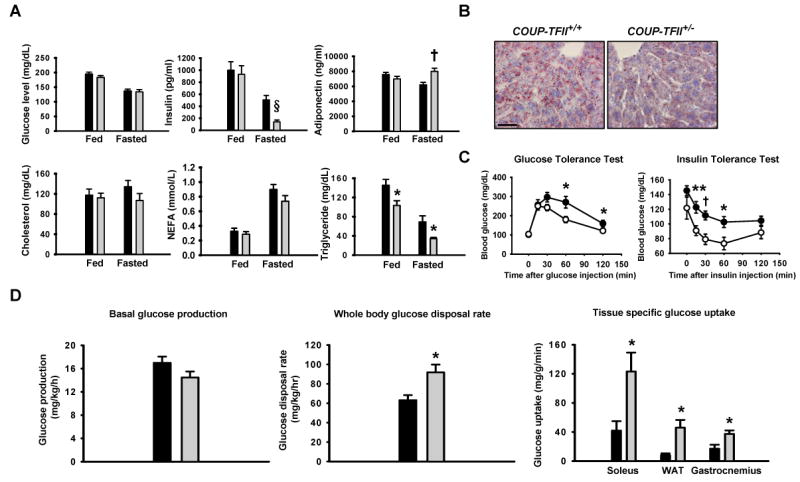
Adipose tissue development and function play a central role in the pathogenesis and pathophysiology of metabolic syndromes. Here, we show that chicken ovalbumin upstream promoter transcription factor II (COUP-TFII) plays a pivotal role in adipogenesis and energy homeostasis. COUP-TFII is expressed in the early stages of white adipocyte development. COUP-TFII heterozygous mice (COUP-TFII(+/-)) have much less white adipose tissue (WAT) than wild-type mice (COUP-TFII(+/+)). COUP-TFII(+/-) mice display a decreased expression of key regulators for WAT development. Knockdown COUP-TFII in 3T3-L1 cells resulted in an increased expression of Wnt10b, while chromatin immunoprecipitation analysis revealed that Wnt10b is a direct target of COUP-TFII. Moreover, COUP-TFII(+/-) mice have increased mitochondrial biogenesis in WAT, and COUP-TFII(+/-) mice have improved glucose homeostasis and increased energy expenditure. Thus, COUP-TFII regulates adipogenesis by regulating the key molecules in adipocyte development and can serve as a target for regulating energy metabolism.
Figures
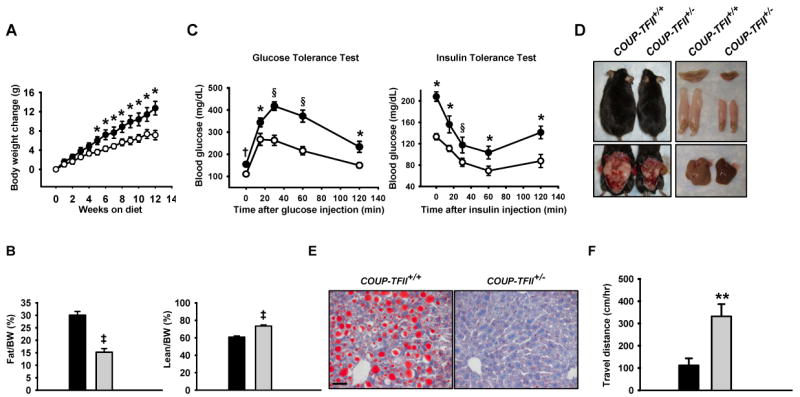
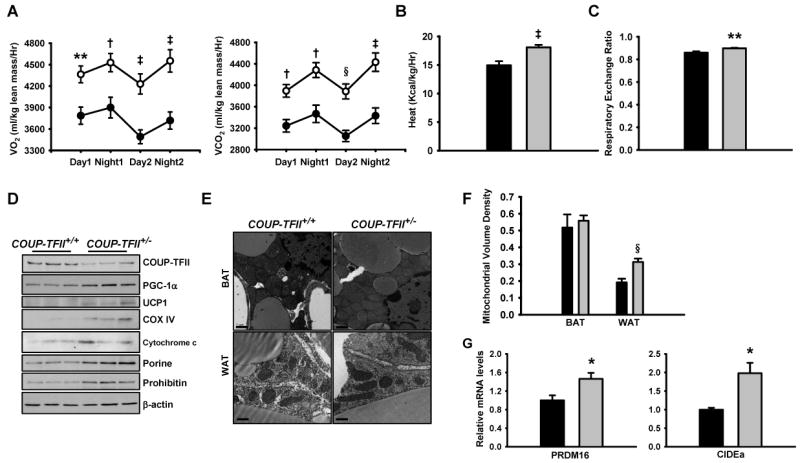
References
-
- Ailhaud G, Grimaldi P, Negrel R. Cellular and molecular aspects of adipose tissue development. Annu Rev Nutr. 1992;12:207–233. - PubMed
-
- Biddinger SB, Kahn CR. From mice to men: insights into the insulin resistance syndromes. Annu Rev Physiol. 2006;68:123–158. - PubMed
-
- Fu M, Sun T, Bookout AL, Downes M, Yu RT, Evans RM, Mangelsdorf DJ. A nuclear receptor atlas: 3T3-L1 adipogenesis. Mol Endocrinol. 2005;19:2437–2450. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R37 HD017379/HD/NICHD NIH HHS/United States
- DK P30 DK079638/DK/NIDDK NIH HHS/United States
- R01 DK045641/DK/NIDDK NIH HHS/United States
- U19 DK062434/DK/NIDDK NIH HHS/United States
- R01 HL076448/HL/NHLBI NIH HHS/United States
- R01 HD017379/HD/NICHD NIH HHS/United States
- R37 DK045641/DK/NIDDK NIH HHS/United States
- P30 DK079638/DK/NIDDK NIH HHS/United States
- P01 HL059314/HL/NHLBI NIH HHS/United States
- R01 HL051586/HL/NHLBI NIH HHS/United States
- HD17379/HD/NICHD NIH HHS/United States
- HL076448/HL/NHLBI NIH HHS/United States
- DK45641/DK/NIDDK NIH HHS/United States
- HL51586/HL/NHLBI NIH HHS/United States
- P01 DK059820/DK/NIDDK NIH HHS/United States
- P01DK59820/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
