

Quantitative assessment of the distributions of membrane conductances involved in action potential backpropagation along basal dendrites
- PMID: 19118105
- PMCID: PMC2666409
- DOI: 10.1152/jn.00651.2007
Quantitative assessment of the distributions of membrane conductances involved in action potential backpropagation along basal dendrites
Abstract
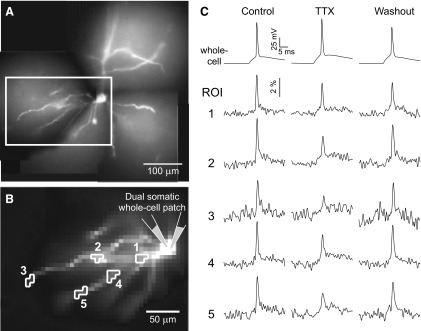
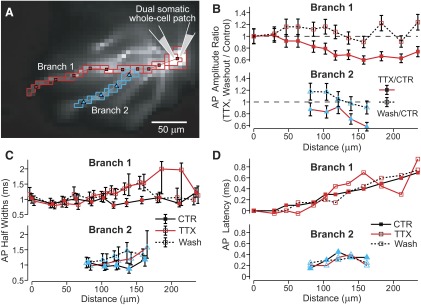
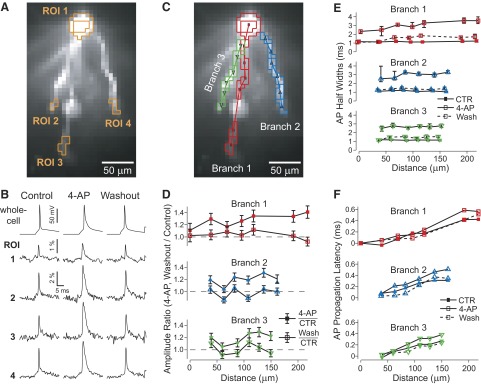
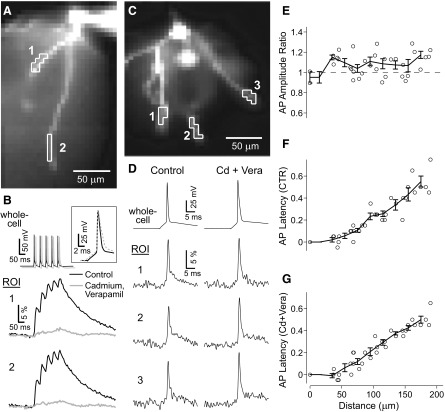
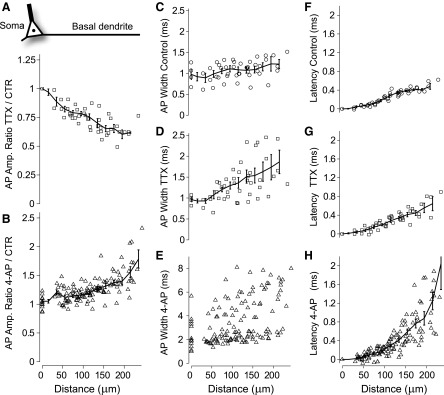
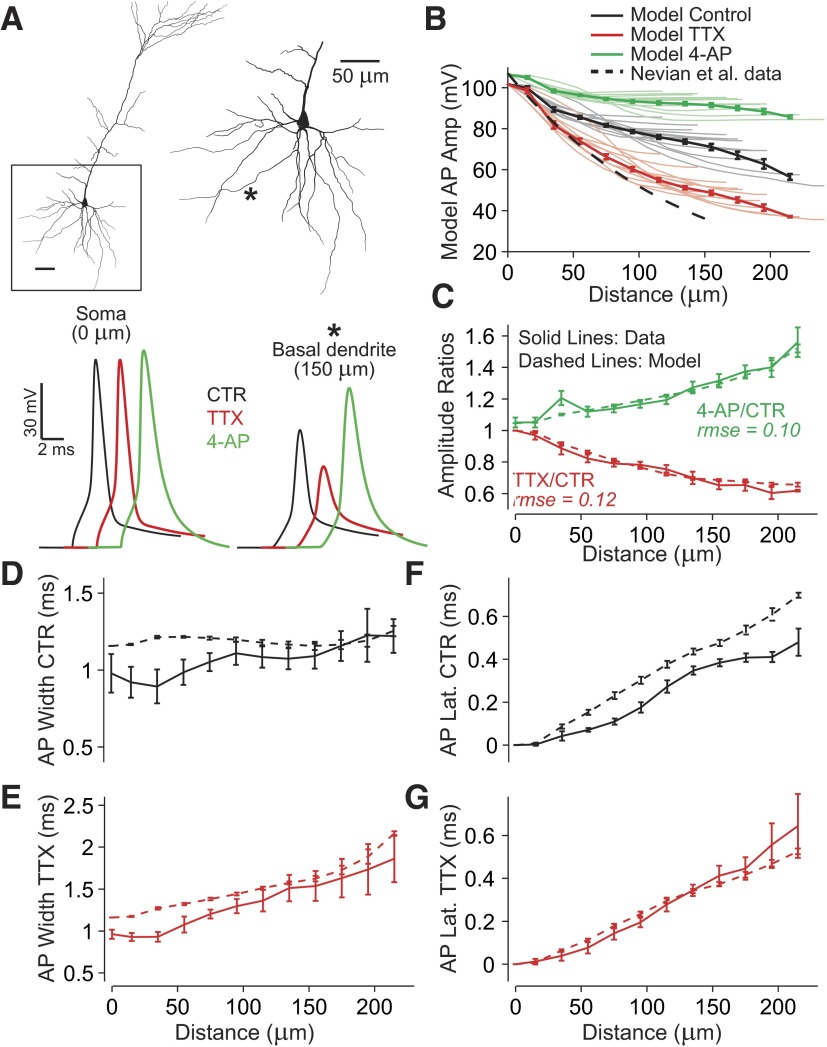
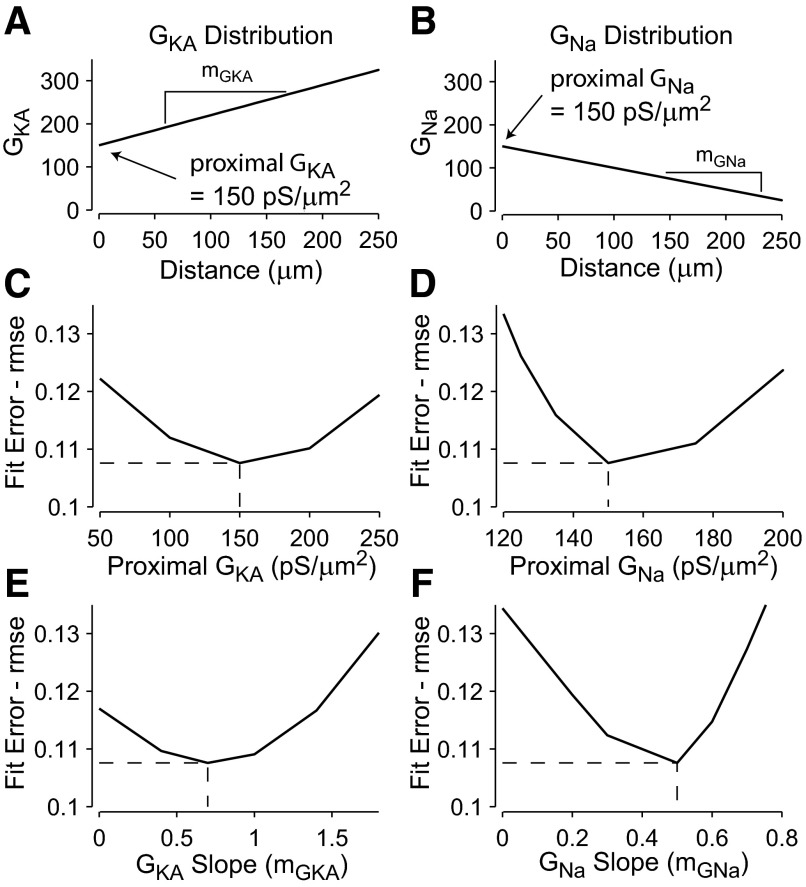
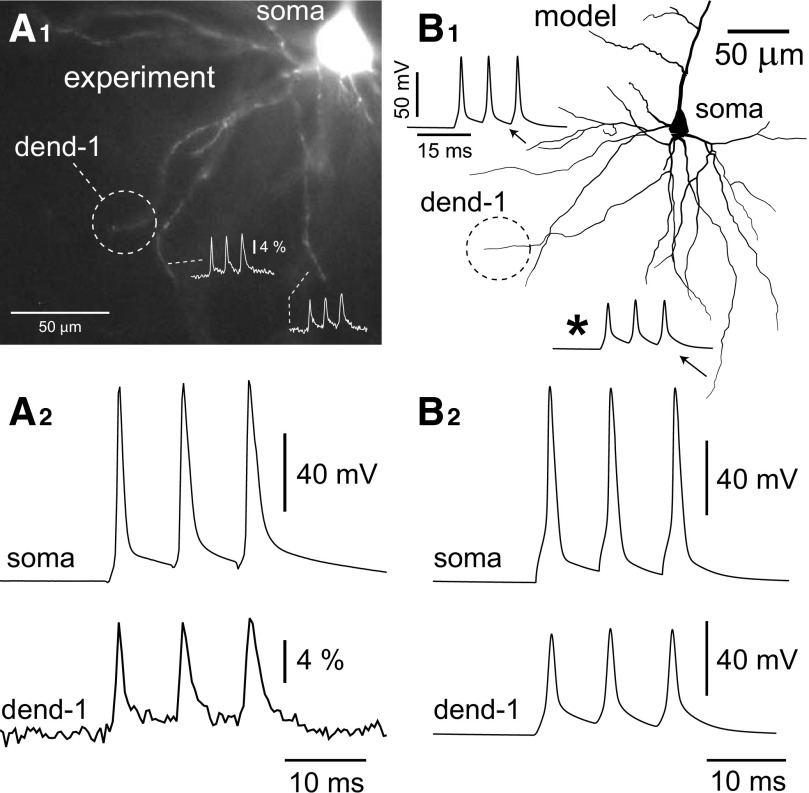
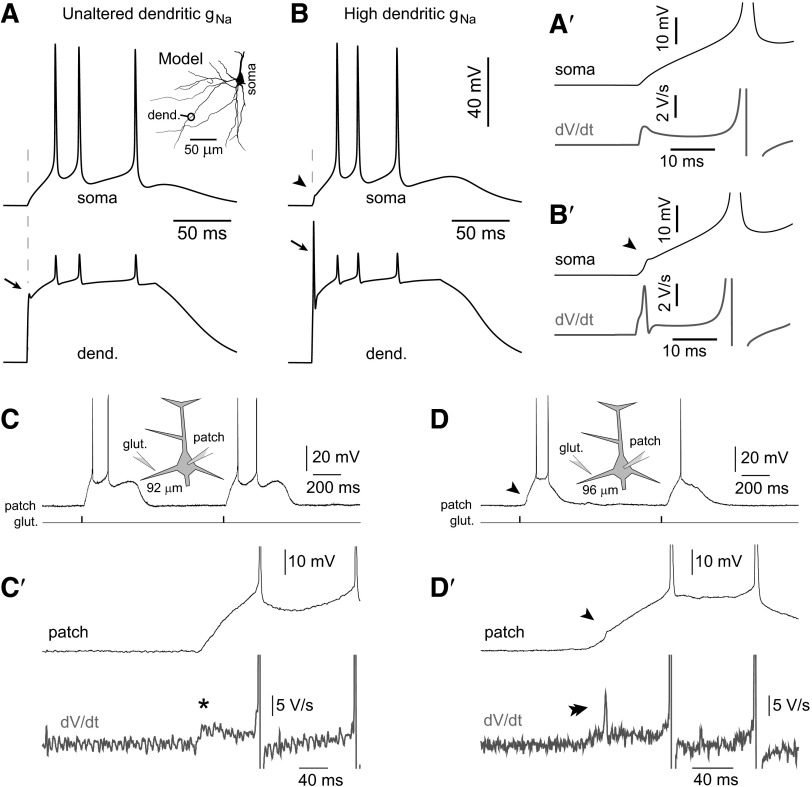
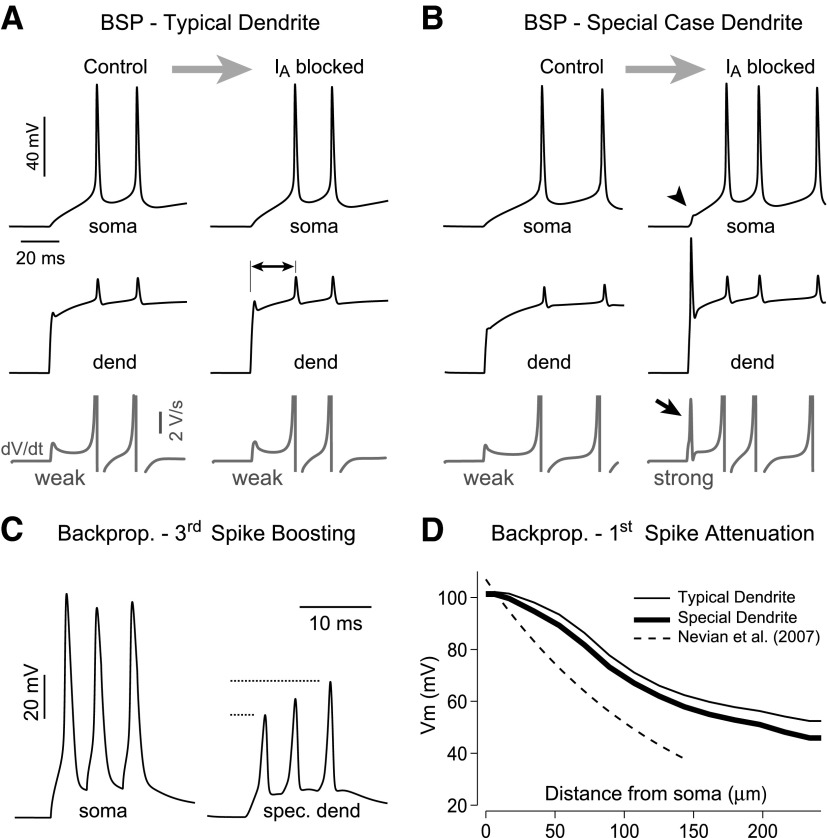
Basal dendrites of prefrontal cortical neurons receive strong synaptic drive from recurrent excitatory synaptic inputs. Synaptic integration within basal dendrites is therefore likely to play an important role in cortical information processing. Both synaptic integration and synaptic plasticity depend crucially on dendritic membrane excitability and the backpropagation of action potentials. We carried out multisite voltage-sensitive dye imaging of membrane potential transients from thin basal branches of prefrontal cortical pyramidal neurons before and after application of channel blockers. We found that backpropagating action potentials (bAPs) are predominantly controlled by voltage-gated sodium and A-type potassium channels. In contrast, pharmacologically blocking the delayed rectifier potassium, voltage-gated calcium, or I(h) conductance had little effect on dendritic AP propagation. Optically recorded bAP waveforms were quantified and multicompartmental modeling was used to link the observed behavior with the underlying biophysical properties. The best-fit model included a nonuniform sodium channel distribution with decreasing conductance with distance from the soma, together with a nonuniform (increasing) A-type potassium conductance. AP amplitudes decline with distance in this model, but to a lesser extent than previously thought. We used this model to explore the mechanisms underlying two sets of published data involving high-frequency trains of APs and the local generation of sodium spikelets. We also explored the conditions under which I(A) down-regulation would produce branch strength potentiation in the proposed model. Finally, we discuss the hypothesis that a fraction of basal branches may have different membrane properties compared with sister branches in the same dendritic tree.
Figures
Similar articles
-
Dendritic voltage-gated ion channels regulate the action potential firing mode of hippocampal CA1 pyramidal neurons.J Neurophysiol. 1999 Oct;82(4):1895-901. doi: 10.1152/jn.1999.82.4.1895. J Neurophysiol. 1999. PMID: 10515978
-
Somatic spikes regulate dendritic signaling in small neurons in the absence of backpropagating action potentials.J Neurosci. 2009 Jun 17;29(24):7803-14. doi: 10.1523/JNEUROSCI.0030-09.2009. J Neurosci. 2009. PMID: 19535592 Free PMC article.
-
Dendritic backpropagation and synaptic plasticity in the mormyrid electrosensory lobe.J Physiol Paris. 2008 Jul-Nov;102(4-6):233-45. doi: 10.1016/j.jphysparis.2008.10.004. Epub 2008 Oct 17. J Physiol Paris. 2008. PMID: 18992811
-
How voltage-gated ion channels alter the functional properties of ganglion and amacrine cell dendrites.Arch Ital Biol. 2002 Oct;140(4):347-59. Arch Ital Biol. 2002. PMID: 12228988 Review.
-
Synaptic integration in dendritic trees.J Neurobiol. 2005 Jul;64(1):75-90. doi: 10.1002/neu.20144. J Neurobiol. 2005. PMID: 15884003 Review.
Cited by
-
Ray Tracing Generalized Tube Primitives: Method and Applications.Eurographics Workshop Vis Comput Biomed. 2019 Jun;38(3):467-478. doi: 10.1111/cgf.13703. Epub 2019 Jul 10. Eurographics Workshop Vis Comput Biomed. 2019. PMID: 31840002 Free PMC article.
-
Location matters: somatic and dendritic SK channels answer to distinct calcium signals.J Neurophysiol. 2015 Jul;114(1):1-5. doi: 10.1152/jn.00181.2014. Epub 2014 Sep 3. J Neurophysiol. 2015. PMID: 25185803 Free PMC article.
-
Basal tree complexity shapes functional pathways in the prefrontal cortex.J Neurophysiol. 2017 Oct 1;118(4):1970-1983. doi: 10.1152/jn.00099.2017. Epub 2017 Jul 12. J Neurophysiol. 2017. PMID: 28701532 Free PMC article.
-
EPSPs Measured in Proximal Dendritic Spines of Cortical Pyramidal Neurons.eNeuro. 2016 May 12;3(2):ENEURO.0050-15.2016. doi: 10.1523/ENEURO.0050-15.2016. eCollection 2016 Mar-Apr. eNeuro. 2016. PMID: 27257618 Free PMC article.
-
Rapid dopaminergic and GABAergic modulation of calcium and voltage transients in dendrites of prefrontal cortex pyramidal neurons.J Physiol. 2012 Aug 15;590(16):3891-911. doi: 10.1113/jphysiol.2011.227157. Epub 2012 May 28. J Physiol. 2012. PMID: 22641784 Free PMC article.
References
-
- Acker CD, White JA. Roles of IA and morphology in action potential propagation in CA1 pyramidal cell dendrites. J Comput Neurosci 23: 201–216, 2007. - PubMed
-
- Antic S, Major G, Zecevic D. Fast optical recordings of membrane potential changes from dendrites of pyramidal neurons. J Neurophysiol 82: 1615–1621, 1999. - PubMed
-
- Bar-Yehuda D, Ben-Porat H, Korngreen A. Dendritic excitability during increased synaptic activity in rat neocortical L5 pyramidal neurons. Eur J Neurosci 28: 2183–2194, 2008. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous