

Picomolar amyloid-beta positively modulates synaptic plasticity and memory in hippocampus
- PMID: 19118188
- PMCID: PMC2673049
- DOI: 10.1523/JNEUROSCI.2692-08.2008
Picomolar amyloid-beta positively modulates synaptic plasticity and memory in hippocampus
Abstract
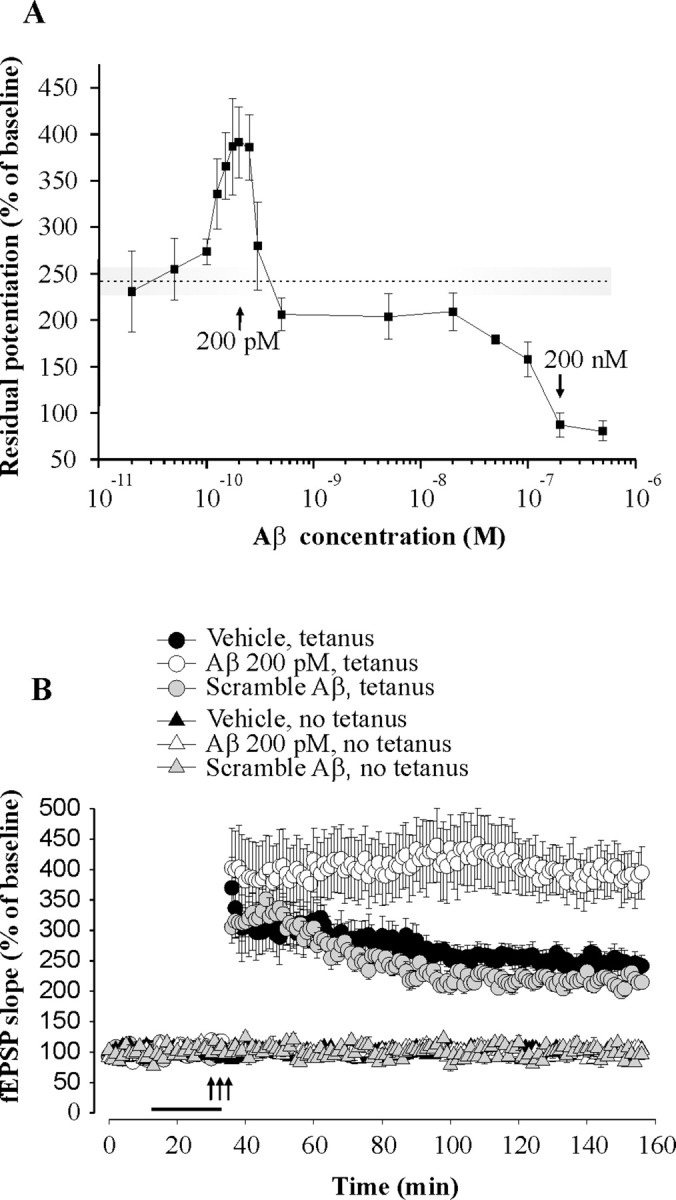
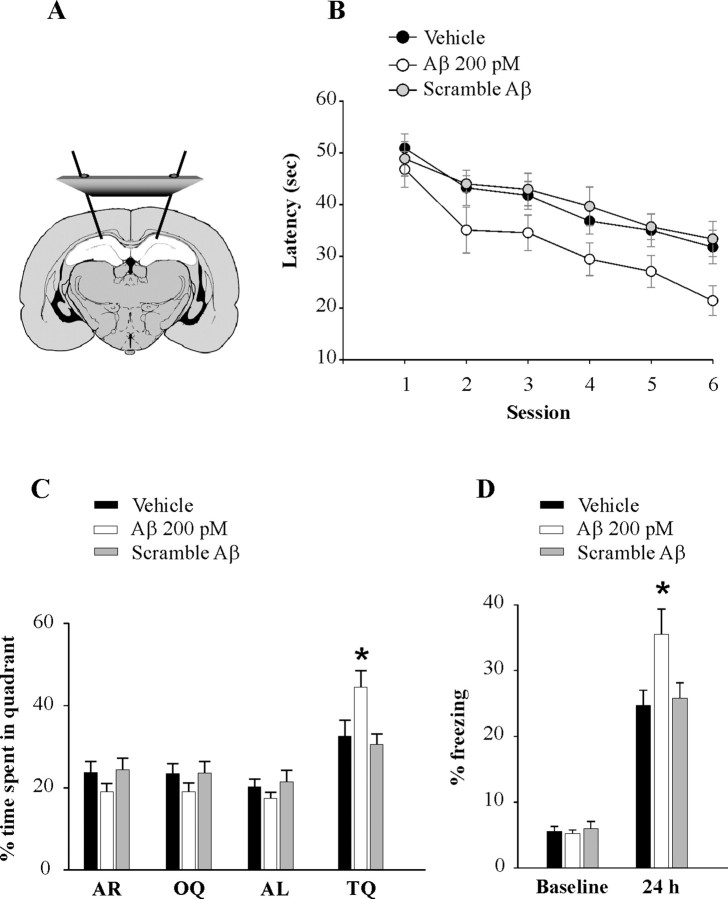
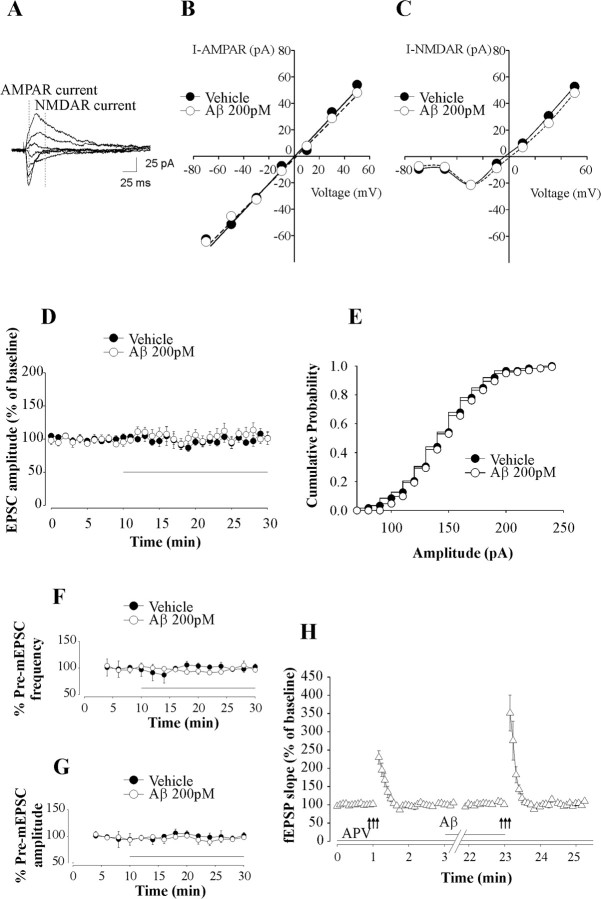
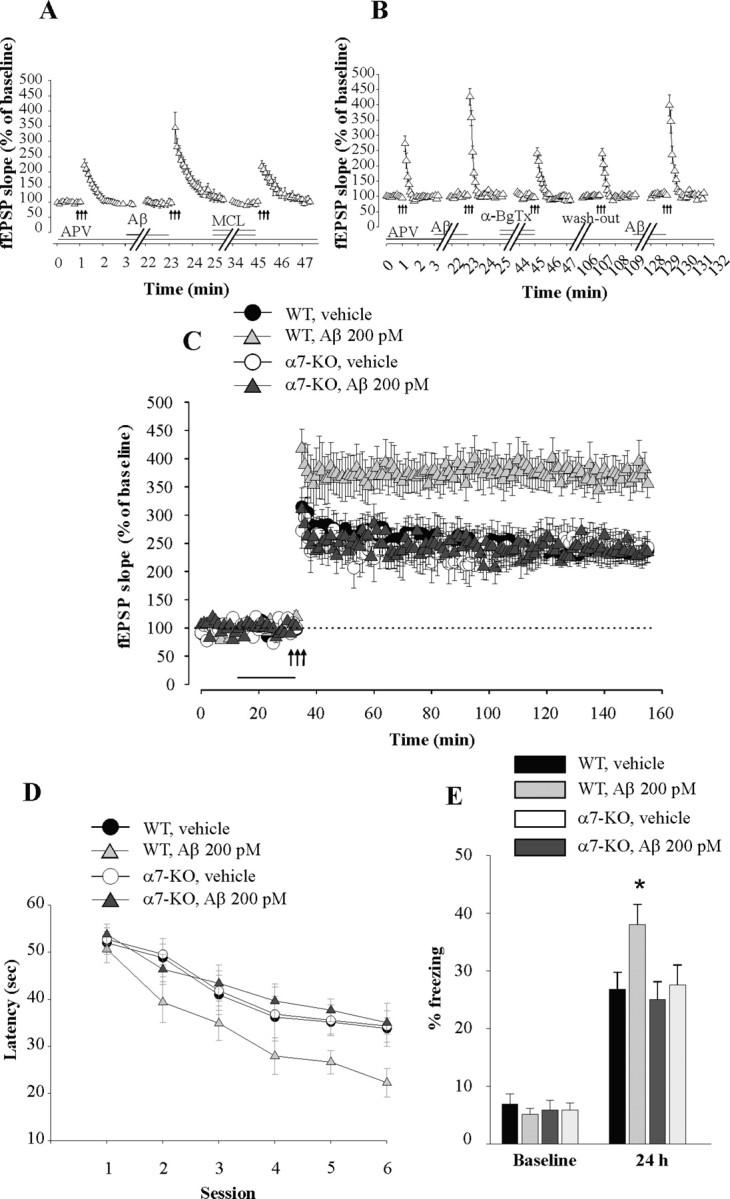
Amyloid-beta (Abeta) peptides are produced in high amounts during Alzheimer's disease, causing synaptic and memory dysfunction. However, they are also released in lower amounts in normal brains throughout life during synaptic activity. Here we show that low picomolar concentrations of a preparation containing both Abeta(42) monomers and oligomers cause a marked increase of hippocampal long-term potentiation, whereas high nanomolar concentrations lead to the well established reduction of potentiation. Picomolar levels of Abeta(42) also produce a pronounced enhancement of both reference and contextual fear memory. The mechanism of action of picomolar Abeta(42) on both synaptic plasticity and memory involves alpha7-containing nicotinic acetylcholine receptors. These findings strongly support a model for Abeta effects in which low concentrations play a novel positive, modulatory role on neurotransmission and memory, whereas high concentrations play the well known detrimental effect culminating in dementia.
Figures
References
-
- ArakiW, Kitaguchi N, Tokushima Y, Ishii K, Aratake H, Shimohama S, Nakamura S, Kimura J (1991) Trophic effect of beta-amyloid precursor protein on cerebral cortical neurons in culture. Biochem Biophys Res Commun 181:265–271. - PubMed
-
- ArancioO, Chao MV (2007) Neurotrophins, synaptic plasticity and dementia. Curr Opin Neurobiol 17:325–330. - PubMed
-
- CaoX, Südhof TC (2001) A transcriptionally [correction of transcriptively] active complex of APP with Fe65 and histone acetyltransferase Tip60. Science 293:115–120. - PubMed
-
- CirritoJR, May PC, O'Dell MA, Taylor JW, Parsadanian M, Cramer JW, Audia JE, Nissen JS, Bales KR, Paul SM, DeMattos RB, Holtzman DM (2003) In vivo assessment of brain interstitial fluid with microdialysis reveals plaque-associated changes in amyloid-β metabolism and half-life. J Neurosci 23:8844–8853. - PMC - PubMed
-
- CirritoJR, Yamada KA, Finn MB, Sloviter RS, Bales KR, May PC, Schoepp DD, Paul SM, Mennerick S, Holtzman DM (2005) Synaptic activity regulates interstitial fluid amyloid-beta levels in vivo. Neuron 48:913–922. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases