

Pyrrolysyl-tRNA synthetase-tRNA(Pyl) structure reveals the molecular basis of orthogonality
- PMID: 19118381
- PMCID: PMC2648862
- DOI: 10.1038/nature07611
Pyrrolysyl-tRNA synthetase-tRNA(Pyl) structure reveals the molecular basis of orthogonality
Abstract
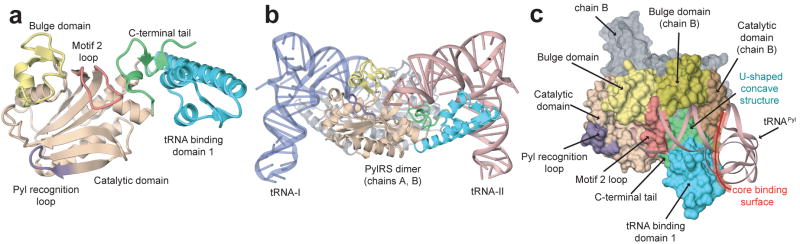
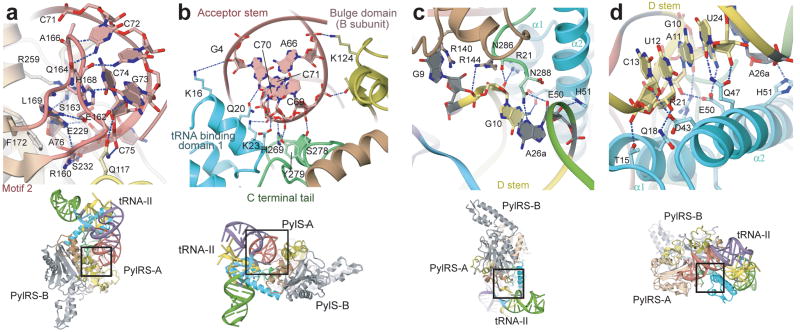
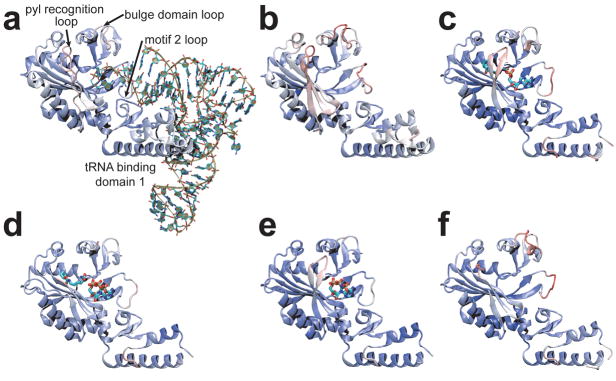
Pyrrolysine (Pyl), the 22nd natural amino acid, is genetically encoded by UAG and inserted into proteins by the unique suppressor tRNA(Pyl) (ref. 1). The Methanosarcinaceae produce Pyl and express Pyl-containing methyltransferases that allow growth on methylamines. Homologous methyltransferases and the Pyl biosynthetic and coding machinery are also found in two bacterial species. Pyl coding is maintained by pyrrolysyl-tRNA synthetase (PylRS), which catalyses the formation of Pyl-tRNA(Pyl) (refs 4, 5). Pyl is not a recent addition to the genetic code. PylRS was already present in the last universal common ancestor; it then persisted in organisms that utilize methylamines as energy sources. Recent protein engineering efforts added non-canonical amino acids to the genetic code. This technology relies on the directed evolution of an 'orthogonal' tRNA synthetase-tRNA pair in which an engineered aminoacyl-tRNA synthetase (aaRS) specifically and exclusively acylates the orthogonal tRNA with a non-canonical amino acid. For Pyl the natural evolutionary process developed such a system some 3 billion years ago. When transformed into Escherichia coli, Methanosarcina barkeri PylRS and tRNA(Pyl) function as an orthogonal pair in vivo. Here we show that Desulfitobacterium hafniense PylRS-tRNA(Pyl) is an orthogonal pair in vitro and in vivo, and present the crystal structure of this orthogonal pair. The ancient emergence of PylRS-tRNA(Pyl) allowed the evolution of unique structural features in both the protein and the tRNA. These structural elements manifest an intricate, specialized aaRS-tRNA interaction surface that is highly distinct from those observed in any other known aaRS-tRNA complex; it is this general property that underlies the molecular basis of orthogonality.
Figures
References
-
- Srinivasan G, James CM, Krzycki JA. Pyrrolysine encoded by UAG in Archaea: charging of a UAG-decoding specialized tRNA. Science. 2002;296:1459–1462. - PubMed
-
- Krzycki JA. The direct genetic encoding of pyrrolysine. Curr Opin Microbiol. 2005;8:706–712. - PubMed
-
- Blight SK, et al. Direct charging of tRNACUA with pyrrolysine in vitro and in vivo. Nature. 2004;431:333–335. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
