

Synthetic control of a fitness tradeoff in yeast nitrogen metabolism
- PMID: 19118500
- PMCID: PMC2631470
- DOI: 10.1186/1754-1611-3-1
Synthetic control of a fitness tradeoff in yeast nitrogen metabolism
Abstract
Background: Microbial communities are involved in many processes relevant to industrial and medical biotechnology, such as the formation of biofilms, lignocellulosic degradation, and hydrogen production. The manipulation of synthetic and natural microbial communities and their underlying ecological parameters, such as fitness, evolvability, and variation, is an increasingly important area of research for synthetic biology.
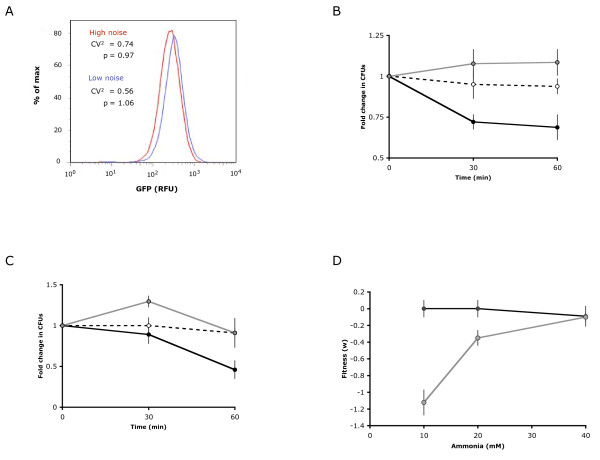
Results: Here, we explored how synthetic control of an endogenous circuit can be used to regulate a tradeoff between fitness in resource abundant and resource limited environments in a population of Saccharomyces cerevisiae. We found that noise in the expression of a key enzyme in ammonia assimilation, Gdh1p, mediated a tradeoff between growth in low nitrogen environments and stress resistance in high ammonia environments. We implemented synthetic control of an endogenous Gdh1p regulatory network to construct an engineered strain in which the fitness of the population was tunable in response to an exogenously-added small molecule across a range of ammonia environments.
Conclusion: The ability to tune fitness and biological tradeoffs will be important components of future efforts to engineer microbial communities.
Figures




Similar articles
-
Hybridization and adaptive evolution of diverse Saccharomyces species for cellulosic biofuel production.Biotechnol Biofuels. 2017 Mar 27;10:78. doi: 10.1186/s13068-017-0763-7. eCollection 2017. Biotechnol Biofuels. 2017. PMID: 28360936 Free PMC article.
-
Engineering Kluyveromyces marxianus as a Robust Synthetic Biology Platform Host.mBio. 2018 Sep 25;9(5):e01410-18. doi: 10.1128/mBio.01410-18. mBio. 2018. PMID: 30254120 Free PMC article.
-
Coupling gene regulatory patterns to bioprocess conditions to optimize synthetic metabolic modules for improved sesquiterpene production in yeast.Biotechnol Biofuels. 2017 Feb 21;10:43. doi: 10.1186/s13068-017-0728-x. eCollection 2017. Biotechnol Biofuels. 2017. PMID: 28239415 Free PMC article.
-
Production of fuels and chemicals from xylose by engineered Saccharomyces cerevisiae: a review and perspective.Microb Cell Fact. 2017 May 11;16(1):82. doi: 10.1186/s12934-017-0694-9. Microb Cell Fact. 2017. PMID: 28494761 Free PMC article. Review.
-
Value-added biotransformation of cellulosic sugars by engineered Saccharomyces cerevisiae.Bioresour Technol. 2018 Jul;260:380-394. doi: 10.1016/j.biortech.2018.04.013. Epub 2018 Apr 7. Bioresour Technol. 2018. PMID: 29655899 Review.
Cited by
-
Mapping the environmental fitness landscape of a synthetic gene circuit.PLoS Comput Biol. 2012;8(4):e1002480. doi: 10.1371/journal.pcbi.1002480. Epub 2012 Apr 12. PLoS Comput Biol. 2012. PMID: 22511863 Free PMC article.
-
Noise minimisation in gene expression switches.PLoS One. 2013 Dec 23;8(12):e84020. doi: 10.1371/journal.pone.0084020. eCollection 2013. PLoS One. 2013. PMID: 24376783 Free PMC article.
-
Network inference and network response identification: moving genome-scale data to the next level of biological discovery.Mol Biosyst. 2010 Mar;6(3):469-80. doi: 10.1039/b916989j. Epub 2009 Dec 11. Mol Biosyst. 2010. PMID: 20174676 Free PMC article. Review.
-
Tuning and controlling gene expression noise in synthetic gene networks.Nucleic Acids Res. 2010 May;38(8):2712-26. doi: 10.1093/nar/gkq091. Epub 2010 Mar 8. Nucleic Acids Res. 2010. PMID: 20211838 Free PMC article.
-
Using design of experiments to guide genetic optimization of engineered metabolic pathways.J Ind Microbiol Biotechnol. 2024 Jan 9;51:kuae010. doi: 10.1093/jimb/kuae010. J Ind Microbiol Biotechnol. 2024. PMID: 38490746 Free PMC article. Review.
References
-
- Flint HJ, Bayer EA. Plant cell wall breakdown by anaerobic microorganisms from the Mammalian digestive tract. Ann N Y Acad Sci. 2008;1125:280–288. - PubMed
-
- Icgen B, Moosa S, Harrison ST. A study of the relative dominance of selected anaerobic sulfate-reducing bacteria in a continuous bioreactor by fluorescence in situ hybridization. Microb Ecol. 2007;53:43–52. - PubMed
-
- Antic MP, Jovancicevic BS, Ilic M, Vrvic MM, Schwarzbauer J. Petroleum pollutant degradation by surface water microorganisms. Environ Sci Pollut Res Int. 2006;13:320–327. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
