

A novel mode of translocation for cytolethal distending toxin
- PMID: 19118582
- PMCID: PMC2647582
- DOI: 10.1016/j.bbamcr.2008.11.017
A novel mode of translocation for cytolethal distending toxin
Abstract
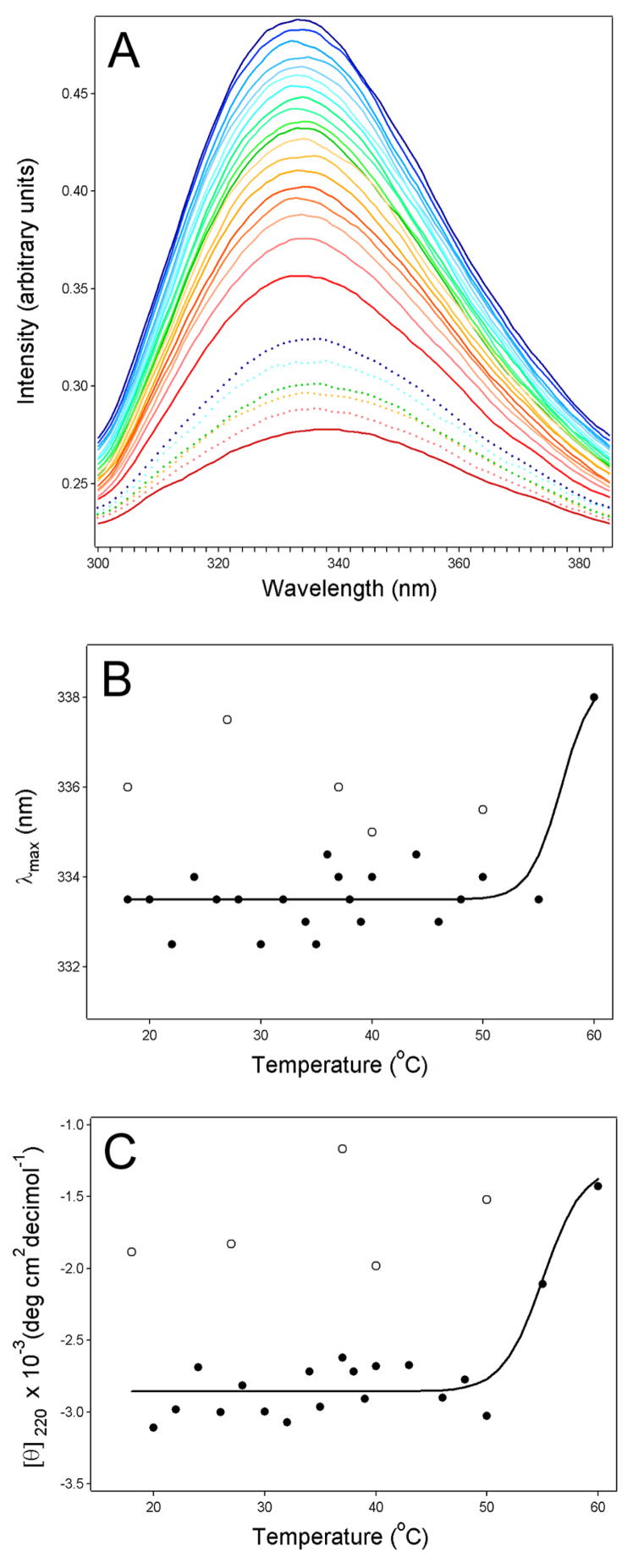
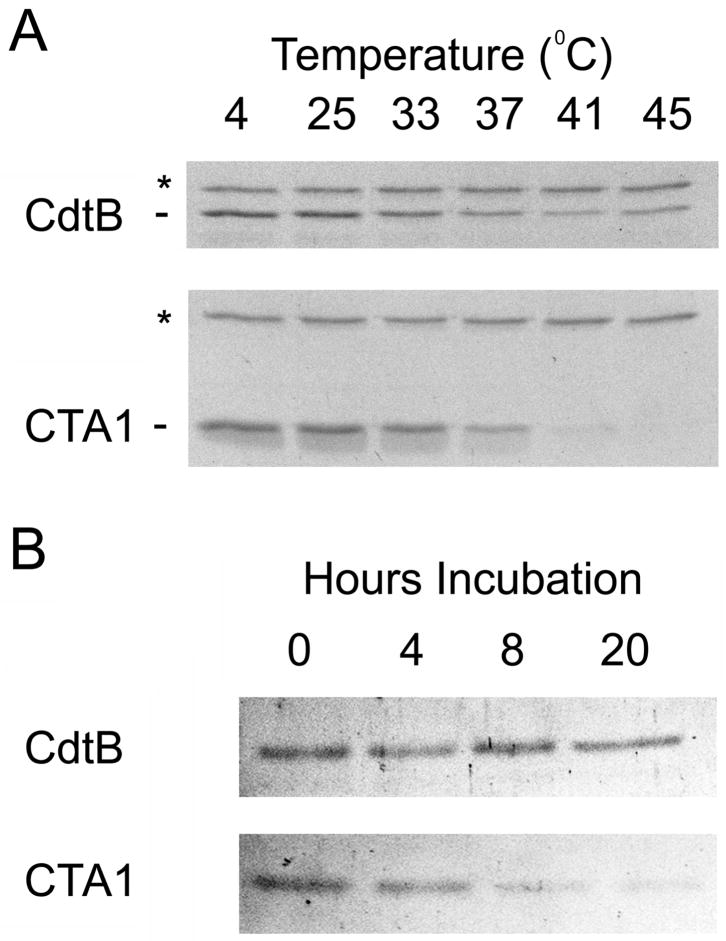
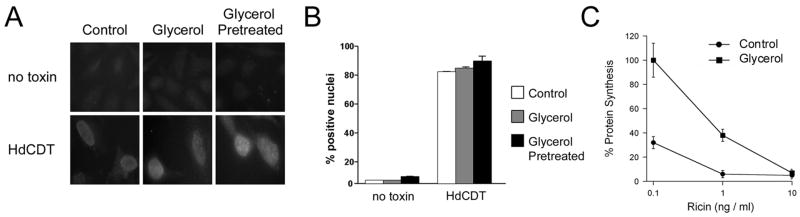
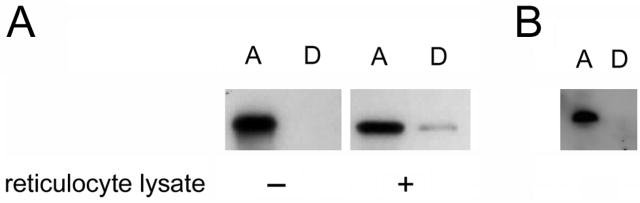
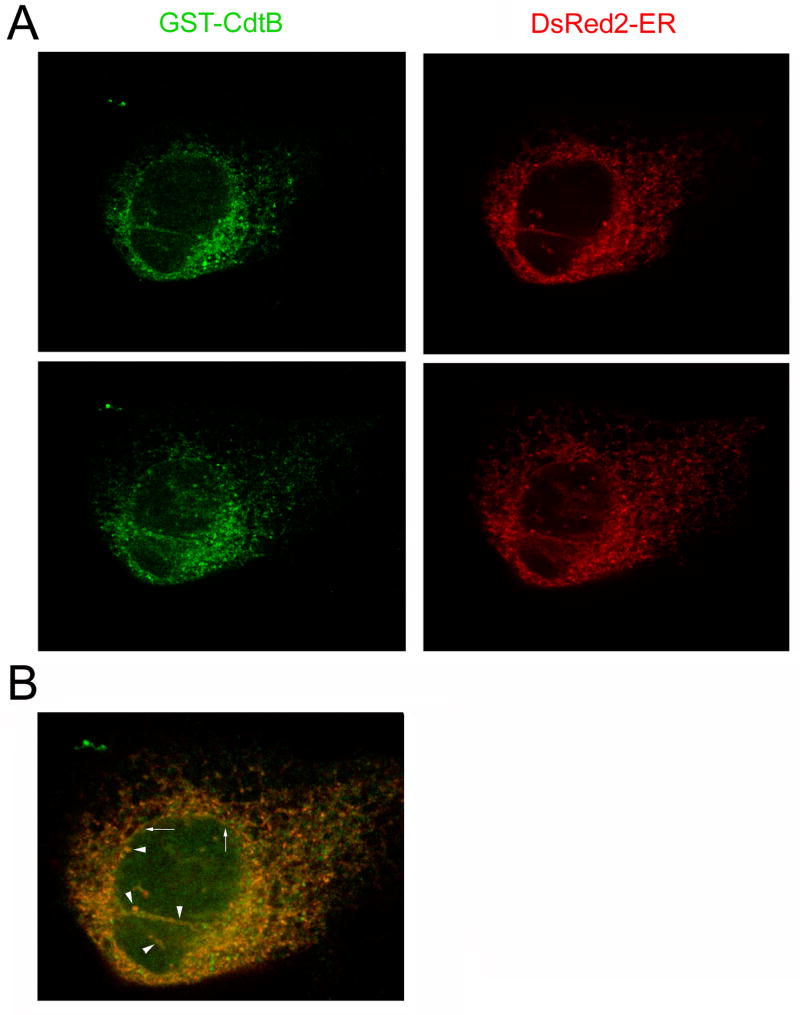
Thermal instability in the toxin catalytic subunit may be a common property of toxins that exit the endoplasmic reticulum (ER) by exploiting the mechanism of ER-associated degradation (ERAD). The Haemophilus ducreyi cytolethal distending toxin (HdCDT) does not utilize ERAD to exit the ER, so we predicted the structural properties of its catalytic subunit (HdCdtB) would differ from other ER-translocating toxins. Here, we document the heat-stable properties of HdCdtB which distinguish it from other ER-translocating toxins. Cell-based assays further suggested that HdCdtB does not unfold before exiting the ER and that it may move directly from the ER lumen to the nucleoplasm. These observations suggest a novel mode of ER exit for HdCdtB.
Figures
References
-
- Thelestam M, Frisan T. Cytolethal distending toxins. In: Alouf JE, Popoff MR, editors. The Comprehensive Sourcebook of Bacterial Protein Toxins. Academic Press; San Diego: 2006. pp. 448–467.
-
- Guerra L, Teter K, Lilley BN, Stenerlow B, Holmes RK, Ploegh HL, Sandvig K, Thelestam M, Frisan T. Cellular internalization of cytolethal distending toxin: a new end to a known pathway. Cell Microbiol. 2005;7:921–934. - PubMed
-
- Hazes B, Read RJ. Accumulating evidence suggests that several AB-toxins subvert the endoplasmic reticulum-associated protein degradation pathway to enter target cells. Biochemistry. 1997;36:11051–11054. - PubMed
-
- Lord JM, Roberts LM, Lencer WI. Entry of protein toxins into mammalian cells by crossing the endoplasmic reticulum membrane: co-opting basic mechanisms of endoplasmic reticulum-associated degradation. Curr Top Microbiol Immunol. 2005;300:149–168. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
