

Mechanistic and kinetic details of catalysis of thiol-disulfide exchange by glutaredoxins and potential mechanisms of regulation
- PMID: 19119916
- PMCID: PMC2842129
- DOI: 10.1089/ars.2008.2291
Mechanistic and kinetic details of catalysis of thiol-disulfide exchange by glutaredoxins and potential mechanisms of regulation
Abstract
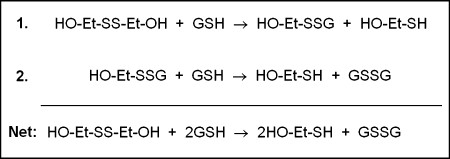
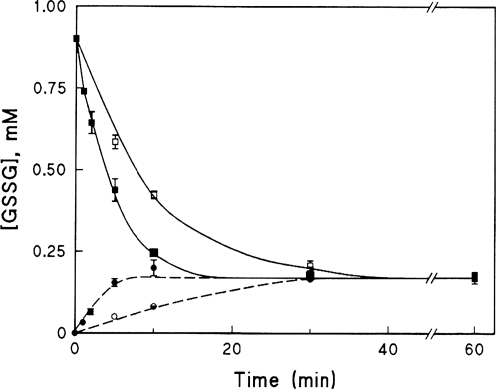
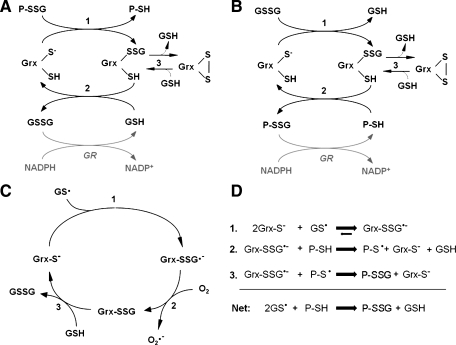
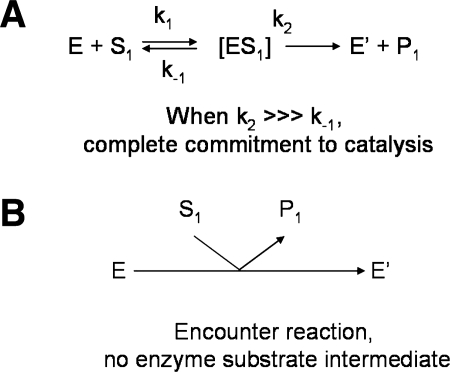
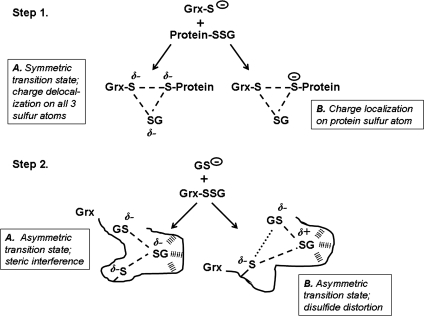
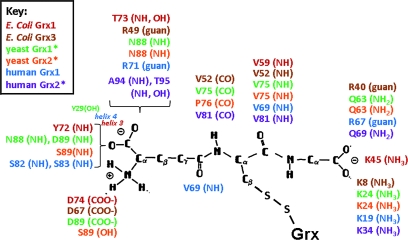
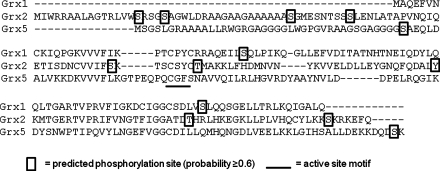
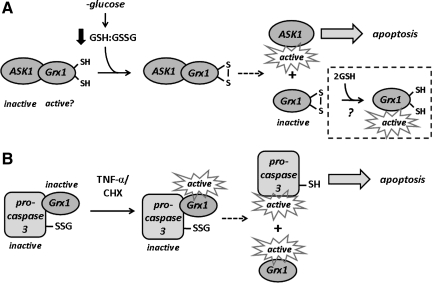
Glutaredoxins are small, heat-stable proteins that exhibit a characteristic thioredoxin fold and a CXXC/S active-site motif. A variety of glutathione (GSH)-dependent catalytic activities have been attributed to the glutaredoxins, including reduction of ribonucleotide reductase, arsenate, and dehydroascorbate; assembly of iron sulfur cluster complexes; and protein glutathionylation and deglutathionylation. Catalysis of reversible protein glutathionylation by glutaredoxins has been implicated in regulation of redox signal transduction and sulfhydryl homeostasis in numerous contexts in health and disease. This forum review is presented in two parts. Part I is focused primarily on the mechanism of the deglutathionylation reaction catalyzed by prototypical dithiol glutaredoxins, especially human Grx1 and Grx2. Grx-catalyzed protein deglutathionylation proceeds by a nucleophilic, double-displacement mechanism in which rate enhancement is attributed to special reactivity of the low pK(a) cysteine at its active site, and to increased nucleophilicity of the second substrate, GSH. Glutaredoxins (and Grx domains) have been identified in most organisms, and many exhibit deglutathionylation or other activities or both. Further characterization according to glutathionyl selectivity, physiological substrates, and intracellular roles may lead to subclassification of this family of enzymes. Part II presents potential mechanisms for in vivo regulation of Grx activity, providing avenues for future studies.
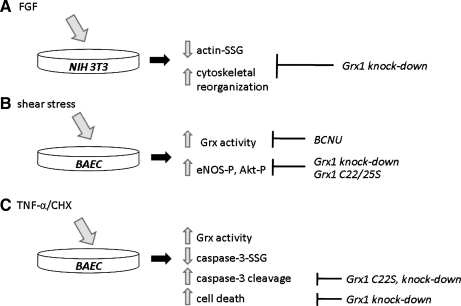
Figures
References
-
- Adachi T. Pimentel DR. Heibeck T. Hou X. Lee YJ. Jiang B. Ido Y. Cohen RA. S-glutathiolation of Ras mediates redox-sensitive signaling by angiotensin II in vascular smooth muscle cells. J Biol Chem. 2004;279:29857–29862. - PubMed
-
- Adachi T. Weisbrod RM. Pimentel DR. Ying J. Sharov VS. Schoneich C. Cohen RA. S-Glutathiolation by peroxynitrite activates SERCA during arterial relaxation by nitric oxide. Nat Med. 2004;10:1200–1207. - PubMed
-
- Askelof P. Axelsson K. Eriksson S. Mannervik B. Mechanism of action of enzymes catalyzing thiol-disulfide interchange: thioltransferases rather than transhydrogenases. FEBS Lett. 1974;38:263–267. - PubMed
-
- Aykac-Toker G. Bulgurcuoglu S. Kocak-Toker N. Effect of peroxynitrite on glutaredoxin. Hum Exp Toxicol. 2001;20:373–376. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
