

A genetically encoded fluorescent reporter of ATP:ADP ratio
- PMID: 19122669
- PMCID: PMC2633436
- DOI: 10.1038/nmeth.1288
A genetically encoded fluorescent reporter of ATP:ADP ratio
Abstract
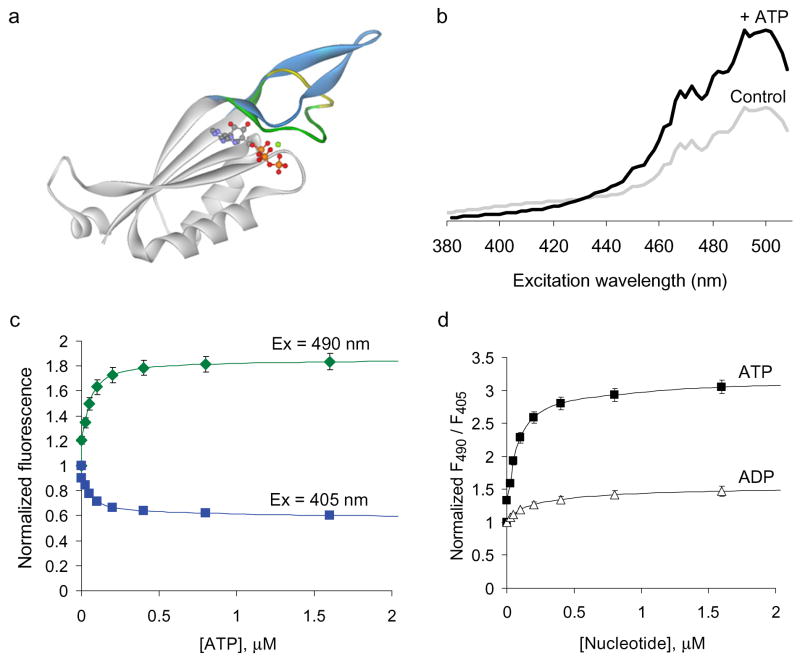
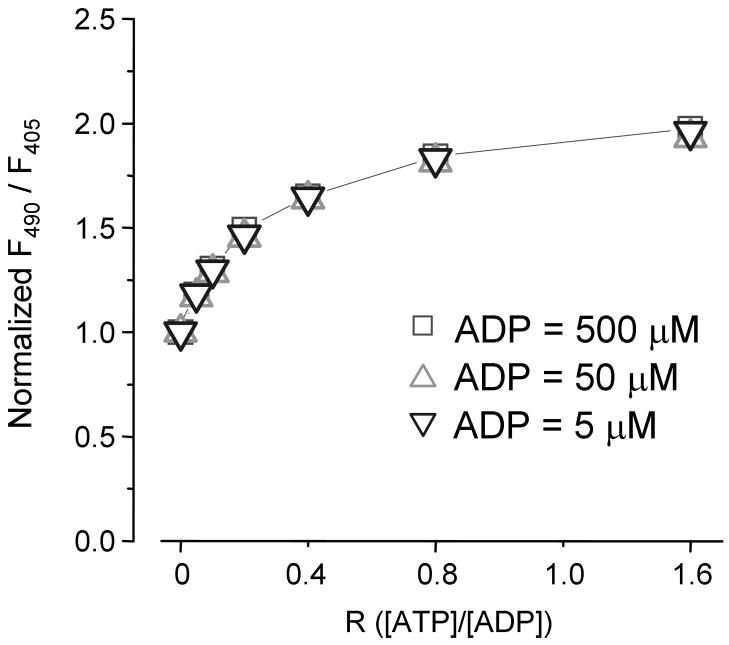
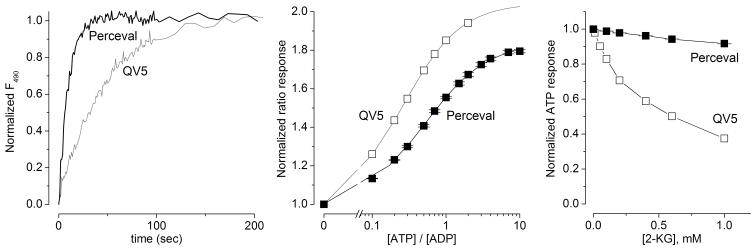
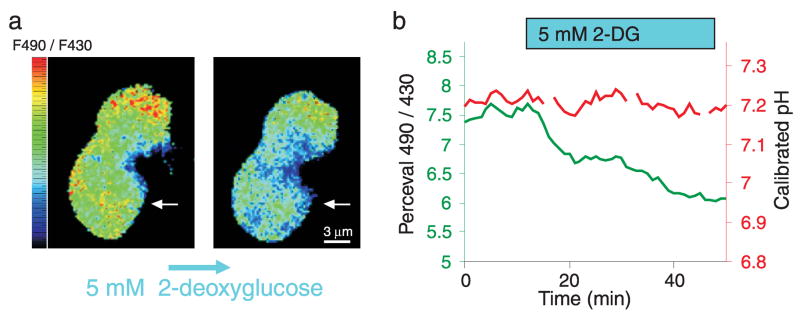
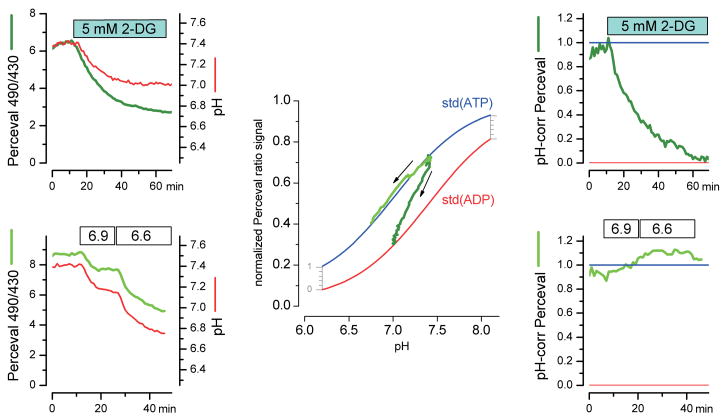
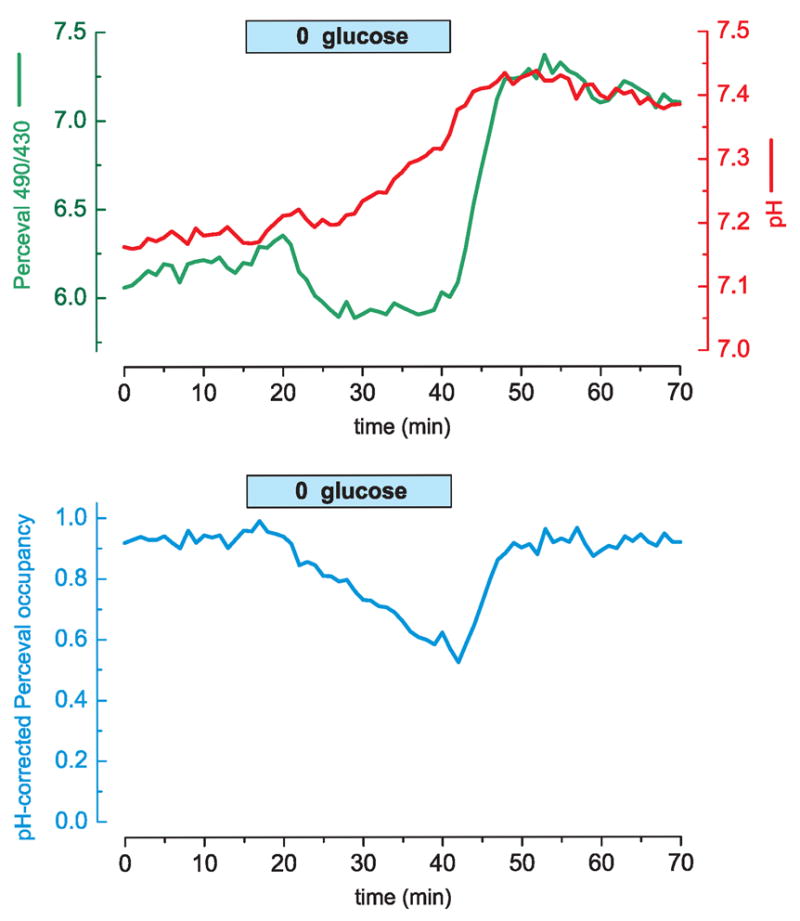
We constructed a fluorescent sensor of adenylate nucleotides by combining a circularly permuted variant of GFP with a bacterial regulatory protein, GlnK1, from Methanococcus jannaschii. The sensor's affinity for Mg-ATP was <100 nM, as seen for other members of the bacterial PII regulator family, a surprisingly high affinity given that normal intracellular ATP concentration is in the millimolar range. ADP bound the same site of the sensor as Mg-ATP, competing with it, but produced a smaller change in fluorescence. At physiological ATP and ADP concentrations, the binding site is saturated, but competition between the two substrates causes the sensor to behave as a nearly ideal reporter of the ATP:ADP concentration ratio. This principle for sensing the ratio of two analytes by competition at a high-affinity site probably underlies the normal functioning of PII regulatory proteins. The engineered sensor, Perceval, can be used to monitor the ATP:ADP ratio during live-cell imaging.
Figures
References
-
- Ashcroft FM, Gribble FM. ATP-sensitive K+ channels and insulin secretion: their role in health and disease. Diabetologia. 1999;42:903–919. - PubMed
-
- Dennis PB, Jaeschke A, Saitoh M, Fowler B, Kozma SC, Thomas G. Mammalian TOR: a homeostatic ATP sensor. Science. 2001;294:1102–1105. - PubMed
-
- Hoffman JF. ATP compartmentation in human erythrocytes. Curr Opin Hematol. 1997;4:112–115. - PubMed
-
- Wilson T, Hastings JW. Bioluminescence. Annu Rev Cell Dev Biol. 1998;14:197–230. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
