

Tricks with tetramers: how to get the most from multimeric peptide-MHC
- PMID: 19125886
- PMCID: PMC2632693
- DOI: 10.1111/j.1365-2567.2008.02848.x
Tricks with tetramers: how to get the most from multimeric peptide-MHC
Erratum in
- Immunology. 2009 Mar;126(3):447
Abstract
The development of fluorochrome-conjugated peptide-major histocompatibility complex (pMHC) multimers in conjunction with continuing advances in flow cytometry has transformed the study of antigen-specific T cells by enabling their visualization, enumeration, phenotypic characterization and isolation from ex vivo samples. Here, we bring together and discuss some of the 'tricks' that can be used to get the most out of pMHC multimers. These include: (1) simple procedures that can substantially enhance the staining intensity of cognate T cells with pMHC multimers; (2) the use of pMHC multimers to stain T cells with very-low-affinity T-cell receptor (TCR)/pMHC interactions, such as those that typically predominate in tumour-specific responses; and (3) the physical grading and clonotypic dissection of antigen-specific T cells based on the affinity of their cognate TCR using mutant pMHC multimers in conjunction with new approaches to the molecular analysis of TCR gene expression. We also examine how soluble pMHC can be used to examine T-cell activation, manipulate T-cell responses and study allogeneic and superantigen interactions with TCRs. Finally, we discuss the problems that arise with pMHC class II (pMHCII) multimers because of the low affinity of TCR/pMHCII interactions and lack of 'coreceptor help'.
Figures
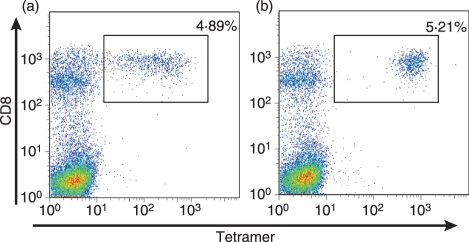
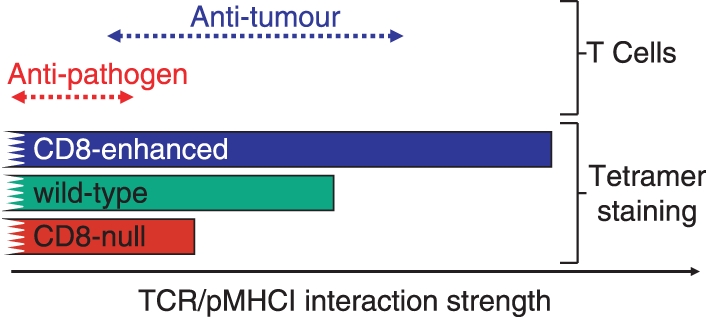
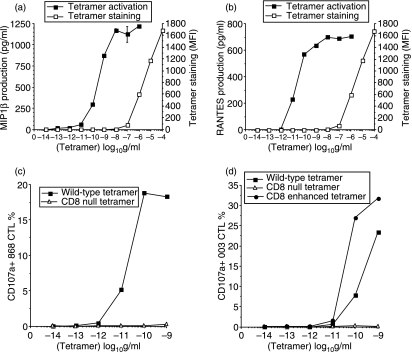
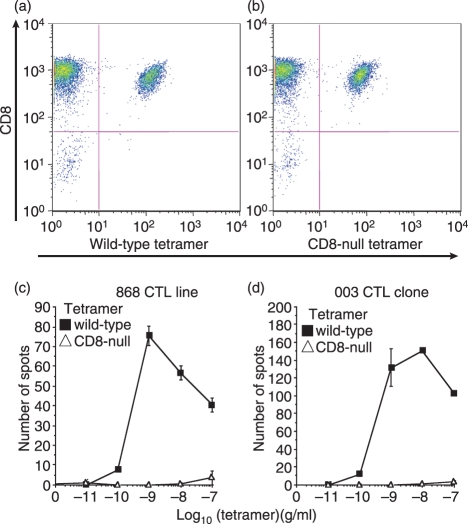
References
-
- Laugel B, Boulter JM, Lissin N, et al. Design of soluble recombinant T cell receptors for antigen targeting and T cell inhibition. J Biol Chem. 2005;280:1882–92. - PubMed
-
- Altman JD, Moss PA, Goulder PJ, Barouch DH, McHeyzer-Williams MG, Bell JI, McMichael AJ, Davis MM. Phenotypic analysis of antigen-specific T lymphocytes. Science. 1996;274:94–6. - PubMed
-
- Guillaume P, Legler DF, Boucheron N, Doucey MA, Cerottini JC, Luescher IF. Soluble major histocompatibility complex–peptide octamers with impaired CD8 binding selectively induce Fas-dependent apoptosis. J Biol Chem. 2003;278:4500–9. - PubMed
-
- Bakker AH, Schumacher TN. MHC multimer technology: current status and future prospects. Curr Opin Immunol. 2005;17:428–33. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
