

Curvature dynamics of alpha-synuclein familial Parkinson disease mutants: molecular simulations of the micelle- and bilayer-bound forms
- PMID: 19126542
- PMCID: PMC2652287
- DOI: 10.1074/jbc.M808895200
Curvature dynamics of alpha-synuclein familial Parkinson disease mutants: molecular simulations of the micelle- and bilayer-bound forms
Abstract
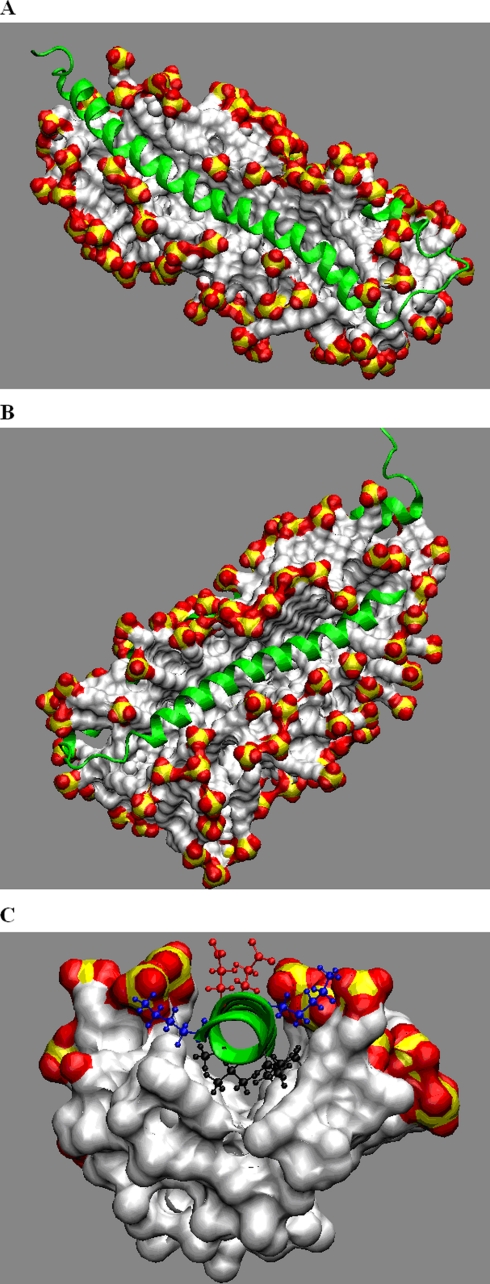
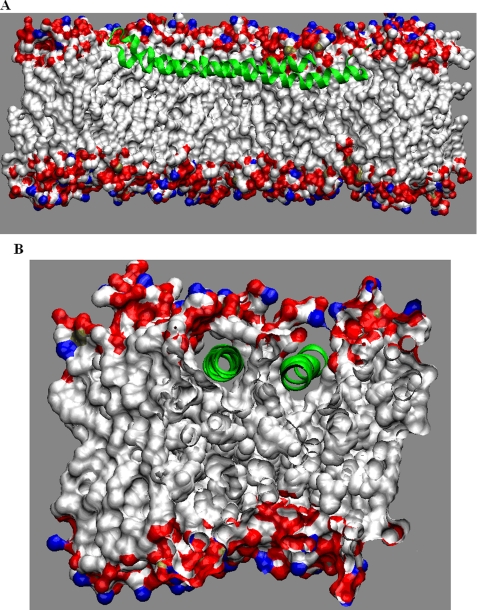
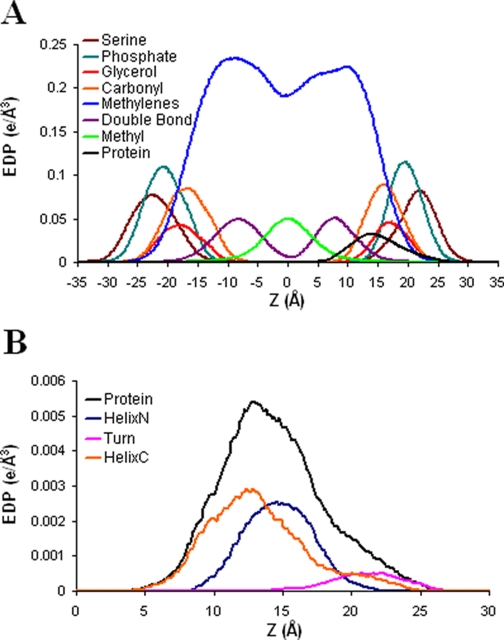
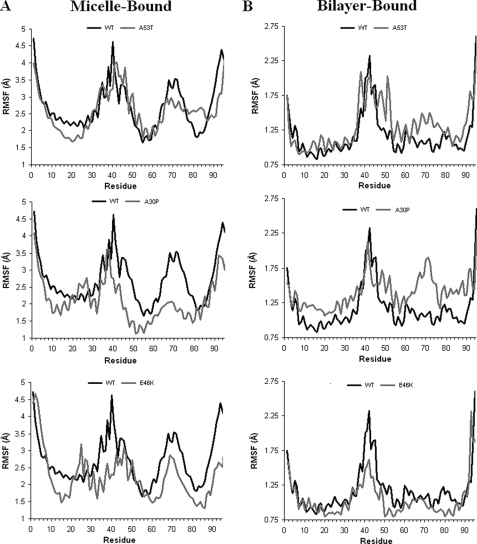
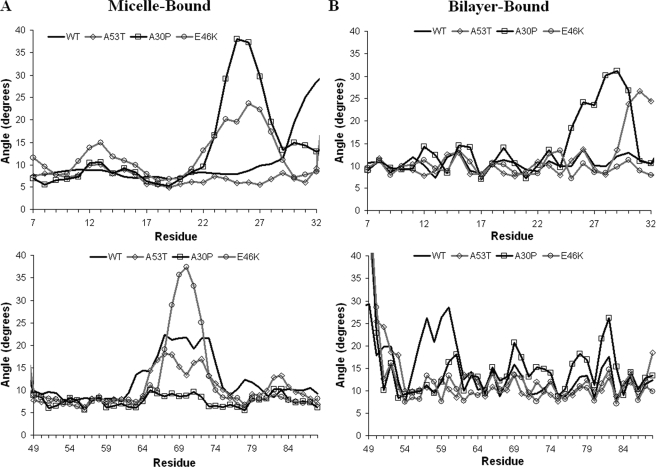
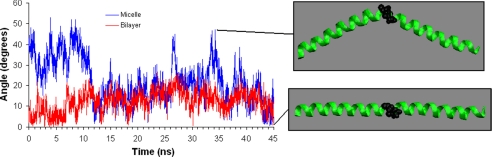
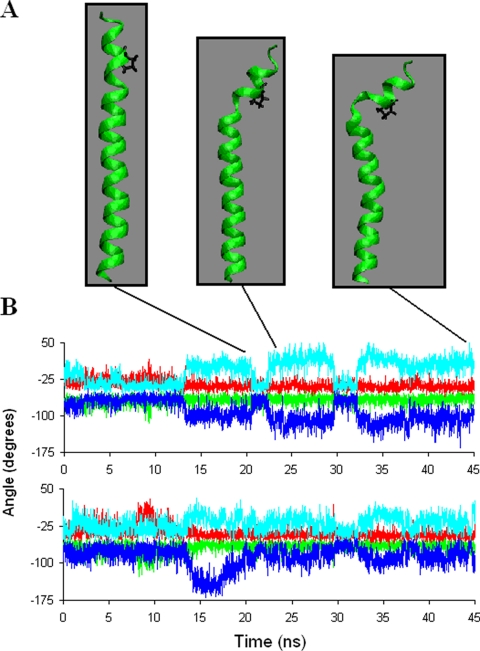
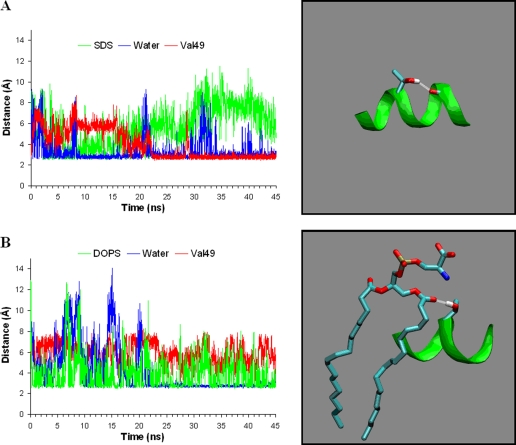
Alpha-synuclein remains a protein of interest due to its propensity to form fibrillar aggregates in neurodegenerative disease and its putative function in synaptic vesicle regulation. Herein, we present a series of atomistic molecular dynamics simulations of wild-type alpha-synuclein and three Parkinson disease familial mutants (A30P, A53T, and E46K) in two distinct environments. First, in order to match recent NMR experiments, we have simulated each protein bound to an SDS detergent micelle. Second, in order to connect more closely to the true biological environment, we have simulated the proteins bound to a 1,2-dioleoyl-sn-glycero-3-phosphoserine lipid bilayer. In the micelle-bound case, we find that the wild type and all of the variants of alpha-synuclein flatten the underlying micelle, decreasing its surface area. A30P is known to lessen alpha-synuclein/membrane affinity and, consistent with experiment, destabilizes the simulated secondary structure. In the case of A53T, our simulations reveal a range of stabilizing hydrogen bonds that form with the threonine. In both environments, the E46K mutation, which is known to increase bilayer affinity, leads to an additional hydrogen bond between the protein and either the detergent or lipid. Simulations indicate that alphaS and its variants are less dynamic in the bilayer than in the micelle. Furthermore, the simulations of the mutants suggest how changes in the structure and dynamics of alpha-synuclein may affect its biological role.
Figures

References
-
- Kruger, R., Kuhn, W., Muller, T., Woitalla, D., Graeber, M., Kosel, S., Przuntek, H., Epplen, J. T., Schols, L., and Riess, O. (1998) Nat. Genet. 18 106-108 - PubMed
-
- Polymeropoulos, M. H., Lavedan, C., Leroy, E., Ide, S. E., Dehejia, A., Dutra, A., Pike, B., Root, H., Rubenstein, J., Boyer, R., Stenroos, E. S., Chandrasekharappa, S., Athanassiadou, A., Papapetropoulos, T., Johnson, W. G., Lazzarini, A. M., Duvoisin, R. C., Di Iorio, G., Golbe, L. I., and Nussbaum, R. L. (1997) Science 276 2045-2047 - PubMed
-
- Zarranz, J. J., Alegre, J., Gomez-Esteban, J. C., Lezcano, E., Ros, R., Ampuero, I., Vidal, L., Hoenicka, J., Rodriguez, O., Atares, B., Llorens, V., Gomez Tortosa, E., del Ser, T., Munoz, D. G., and de Yebenes, J. G. (2004) Ann. Neurol. 55 164-173 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
