

Essential role of Rac1 and Rac3 GTPases in neuronal development
- PMID: 19126596
- PMCID: PMC7617009
- DOI: 10.1096/fj.08-121574
Essential role of Rac1 and Rac3 GTPases in neuronal development
Abstract
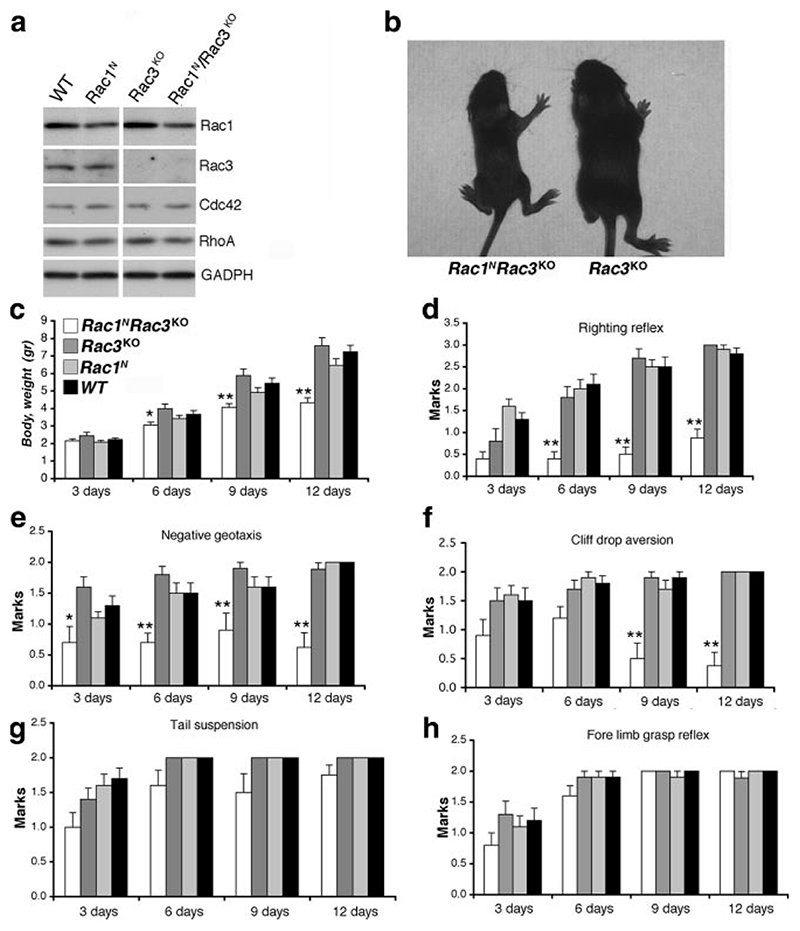
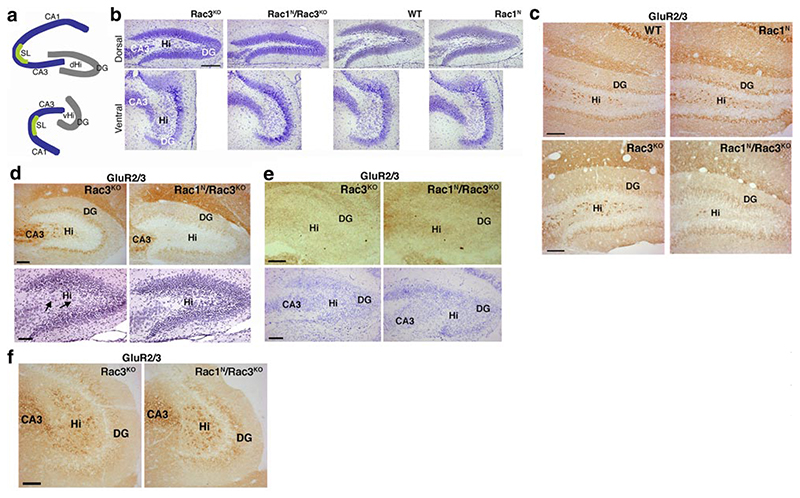
Rac GTPases are members of the Rho family regulating the actin cytoskeleton and implicated in neuronal development. Ubiquitous Rac1 and neuron-specific Rac3 GTPases are coexpressed in the developing mammalian brain. We used Cre-mediated conditional deletion of Rac1 in neurons combined with knockout of neuron-specific Rac3 to study the role of these GTPases in neural development. We found that lack of both genes causes motor behavioral defects, epilepsy, and premature death of mice. Deletion of either GTPase does not produce evident phenotypes. Double-knockout mice show specific defects in the development of the hippocampus. Selective impairment of the dorsal hilus of double-knockout animals is associated with alteration in the formation of the hippocampal circuitry. Axonal pathways to and from the dorsal hilus are affected because of the deficit of hilar mossy cells. Moreover, analysis of Rac function in hippocampal cultures shows that spine formation is strongly hampered only in neurons lacking both Rac proteins. These findings show for the first time that both Rac1 and Rac3 are important for the development of the nervous system, wherein they play complementary roles during late stages of neuronal and brain development.
Figures








References
-
- Hall A. Rho GTPases and the actin cytoskeleton. Science. 1998;279:509–514. - PubMed
-
- Tada T, Sheng M. Molecular mechanisms of dendritic spine morphogenesis. Curr Opin Neurobiol. 2006;16:95–101. - PubMed
-
- De Curtis I. Functions of Rac GTPases during neuronal development. Dev Neurosci. 2008;30:47–58. - PubMed
-
- Didsbury J, Weber RF, Bokoch GM, Evans T, Snyderman R. Rac, a novel ras-related family of proteins that are botulinum toxin substrates. J Biol Chem. 1989;264:16378–16382. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
