

Modification of surfactant protein D by reactive oxygen-nitrogen intermediates is accompanied by loss of aggregating activity, in vitro and in vivo
- PMID: 19126597
- PMCID: PMC2669423
- DOI: 10.1096/fj.08-120568
Modification of surfactant protein D by reactive oxygen-nitrogen intermediates is accompanied by loss of aggregating activity, in vitro and in vivo
Abstract
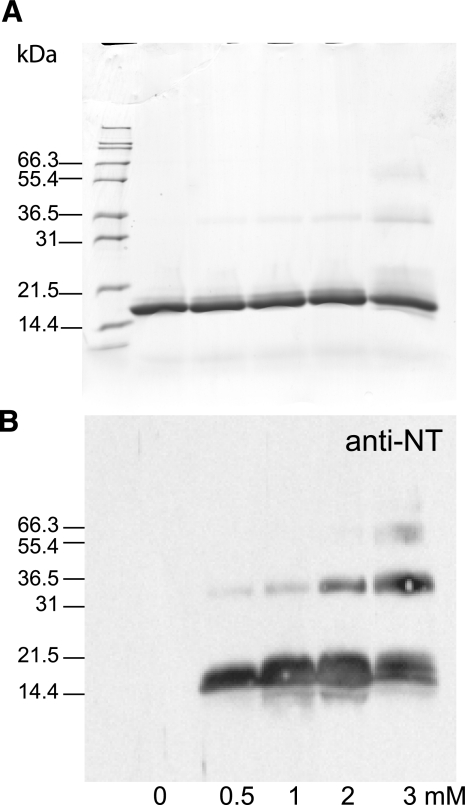
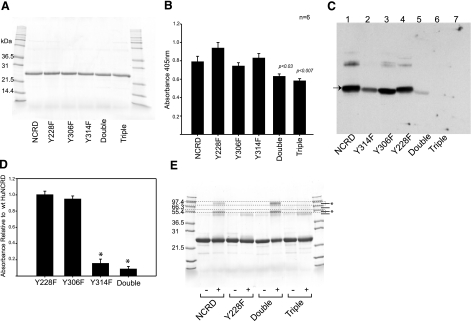
Surfactant protein D (SP-D) is an important effector of innate immunity. We have previously shown that SP-D accumulates at sites of acute bacterial infection and neutrophil infiltration, a setting associated with the release of reactive species such as peroxynitrite. Incubation of native SP-D or trimeric SP-D lectin domains (NCRDs) with peroxynitrite resulted in nitration and nondisulfide cross-linking. Modifications were blocked by peroxynitrite scavengers or pH inactivation of peroxynitrite, and mass spectroscopy confirmed nitration of conserved tyrosine residues within the C-terminal neck and lectin domains. Mutant NCRDs lacking one or more of the tyrosines allowed us to demonstrate preferential nitration of Tyr314 and the formation of Tyr228-dependent cross-links. Although there was no effect of peroxynitrite or tyrosine mutations on lectin activity, incubation of SP-D dodecamers or murine lavage with peroxynitrite decreased the SP-D-dependent aggregation of lipopolysaccharide-coated beads, supporting our hypothesis that defective aggregation results from abnormal cross-linking. We also observed nitration, cross-linking of SP-D, and a significant decrease in SP-D-dependent aggregating activity in the lavage of mice acutely exposed to nitrogen dioxide. Thus, modification of SP-D by reactive oxygen-nitrogen species could contribute to alterations in the structure and function of SP-D at sites of inflammation in vivo.
Figures






References
-
- Kishore U, Greenhough T J, Waters P, Shrive A K, Ghai R, Kamran M F, Bernal A L, Reid K B, Madan T, Chakraborty T. Surfactant proteins SP-A and SP-D: structure, function and receptors. Mol Immunol. 2006;43:1293–1315. - PubMed
-
- Whitsett J A. Surfactant proteins in innate host defense of the lung. Biol Neonate. 2005;88:175–180. - PubMed
-
- Wright J R. Immunoregulatory functions of surfactant proteins. Nat Rev Immunol. 2005;5:58–68. - PubMed
-
- Madsen J, Kliem A, Tornoe I, Skjodt K, Koch C, Holmskov U. Localization of lung surfactant protein D (SP-D) on mucosal surfaces in human tissues. J Immunol. 2000;164:5866–5870. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
