

An endoribonuclease functionally linked to perinuclear mRNP quality control associates with the nuclear pore complexes
- PMID: 19127978
- PMCID: PMC2613419
- DOI: 10.1371/journal.pbio.1000008
An endoribonuclease functionally linked to perinuclear mRNP quality control associates with the nuclear pore complexes
Abstract
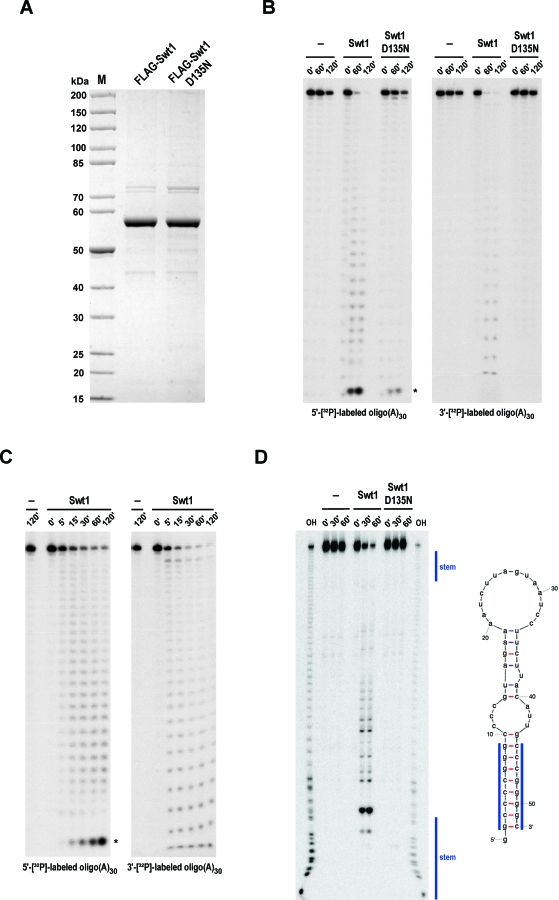
Nuclear mRNA export is a crucial step in eukaryotic gene expression, which is in yeast coupled to cotranscriptional messenger ribonucleoprotein particle (mRNP) assembly and surveillance. Several surveillance systems that monitor nuclear mRNP biogenesis and export have been described, but the mechanism by which the improper mRNPs are recognized and eliminated remains poorly understood. Here we report that the conserved PIN domain protein Swt1 is an RNA endonuclease that participates in quality control of nuclear mRNPs and can associate with the nuclear pore complex (NPC). Swt1 showed endoribonuclease activity in vitro that was inhibited by a point mutation in the predicted catalytic site. Swt1 lacked clear sequence specificity but showed a strong preference for single-stranded regions. Genetic interactions were found between Swt1 and the THO/TREX and TREX-2 complexes, and with components of the perinuclear mRNP surveillance system, Mlp1, Nup60, and Esc1. Inhibition of the nuclease activity of Swt1 increased the levels and cytoplasmic leakage of unspliced aberrant pre-mRNA, and induced robust nuclear poly(A)(+) RNA accumulation in mlp1Delta and esc1Delta strains. Overexpression of Swt1 also caused strong nuclear poly(A)(+) RNA accumulation. Swt1 is normally distributed throughout the nucleus and cytoplasm but becomes concentrated at nuclear pore complexes (NPCs) in the nup133Delta mutant, which causes NPC clustering and defects in mRNP export. The data suggest that Swt1 endoribonuclease might be transiently recruited to NPCs to initiate the degradation of defective pre-mRNPs or mRNPs trapped at nuclear periphery in order to avoid their cytoplasmic export and translation.
Conflict of interest statement
Competing interests. The authors have declared that no competing interests exist.
Figures



Similar articles
-
Nucleocytoplasmic mRNP export is an integral part of mRNP biogenesis.Chromosoma. 2011 Feb;120(1):23-38. doi: 10.1007/s00412-010-0298-1. Epub 2010 Nov 16. Chromosoma. 2011. PMID: 21079985 Free PMC article. Review.
-
Defects in THO/TREX-2 function cause accumulation of novel cytoplasmic mRNP granules that can be cleared by autophagy.RNA. 2016 Aug;22(8):1200-14. doi: 10.1261/rna.057224.116. Epub 2016 Jun 1. RNA. 2016. PMID: 27251550 Free PMC article.
-
A nuclear envelope protein linking nuclear pore basket assembly, SUMO protease regulation, and mRNA surveillance.J Cell Biol. 2007 Aug 27;178(5):813-27. doi: 10.1083/jcb.200702154. J Cell Biol. 2007. PMID: 17724121 Free PMC article.
-
Insights into mRNP biogenesis provided by new genetic interactions among export and transcription factors.BMC Genet. 2012 Sep 10;13:80. doi: 10.1186/1471-2156-13-80. BMC Genet. 2012. PMID: 22963203 Free PMC article.
-
Single particle imaging of mRNAs crossing the nuclear pore: Surfing on the edge.Bioessays. 2016 Aug;38(8):744-50. doi: 10.1002/bies.201600038. Epub 2016 Jun 8. Bioessays. 2016. PMID: 27276446 Review.
Cited by
-
Yeast hEST1A/B (SMG5/6)-like proteins contribute to environment-sensing adaptive gene expression responses.G3 (Bethesda). 2013 Oct 3;3(10):1649-59. doi: 10.1534/g3.113.006924. G3 (Bethesda). 2013. PMID: 23893744 Free PMC article.
-
The nuclear pore complex: bridging nuclear transport and gene regulation.Nat Rev Mol Cell Biol. 2010 Jul;11(7):490-501. doi: 10.1038/nrm2928. Nat Rev Mol Cell Biol. 2010. PMID: 20571586 Review.
-
Regulation of mRNA trafficking by nuclear pore complexes.Genes (Basel). 2014 Sep 2;5(3):767-91. doi: 10.3390/genes5030767. Genes (Basel). 2014. PMID: 25184662 Free PMC article. Review.
-
IMiQ: a novel protein quality control compartment protecting mitochondrial functional integrity.Mol Biol Cell. 2018 Feb 1;29(3):256-269. doi: 10.1091/mbc.E17-01-0027. Epub 2017 Dec 6. Mol Biol Cell. 2018. PMID: 29212875 Free PMC article.
-
mRNA stability in the nucleus.J Zhejiang Univ Sci B. 2014 May;15(5):444-54. doi: 10.1631/jzus.B1400088. J Zhejiang Univ Sci B. 2014. PMID: 24793762 Free PMC article. Review.
References
-
- Köhler A, Hurt E. Exporting RNA from the nucleus to the cytoplasm. Nat Rev Mol Cell Biol. 2007;8:761–773. - PubMed
-
- Rougemaille M, Villa T, Gudipati RK, Libri D. mRNA journey to the cytoplasm: attire required. Biol Cell. 2008;100:327–342. - PubMed
-
- Huertas P, Aguilera A. Cotranscriptionally formed DNA:RNA hybrids mediate transcription elongation impairment and transcription-associated recombination. Mol Cell. 2003;12:711–721. - PubMed
-
- Saguez C, Schmid M, Olesen JR, Ghazy MA, Qu X, et al. Nuclear mRNA surveillance in THO/sub2 mutants is triggered by inefficient polyadenylation. Mol Cell. 2008;31:91–103. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
