

Gene expression studies in kiwifruit and gene over-expression in Arabidopsis indicates that GDP-L-galactose guanyltransferase is a major control point of vitamin C biosynthesis
- PMID: 19129165
- PMCID: PMC2652059
- DOI: 10.1093/jxb/ern327
Gene expression studies in kiwifruit and gene over-expression in Arabidopsis indicates that GDP-L-galactose guanyltransferase is a major control point of vitamin C biosynthesis
Abstract
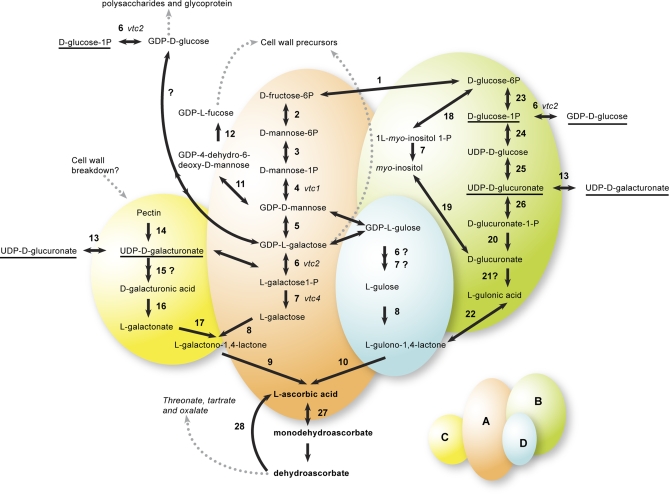
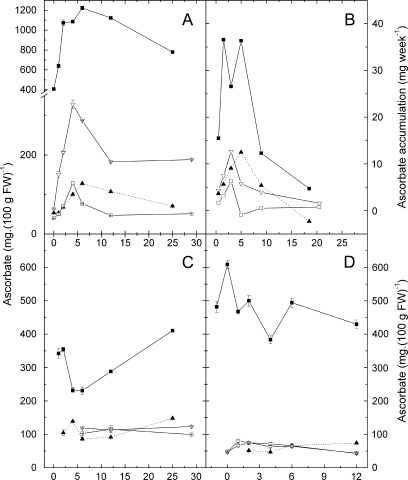
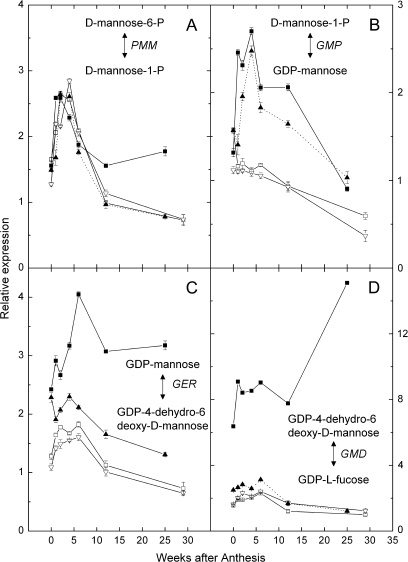
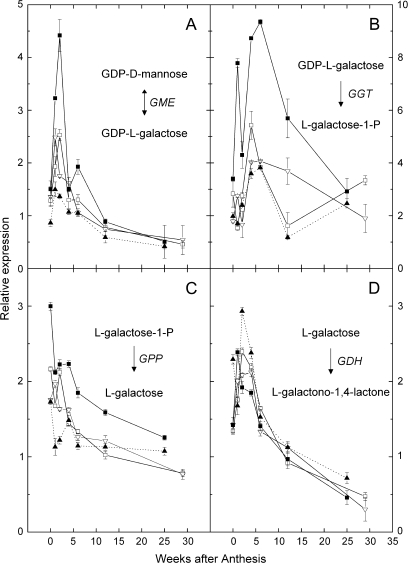
Vitamin C (L-ascorbic acid, AsA) is an essential metabolite for plants and animals. Kiwifruit (Actinidia spp.) are a rich dietary source of AsA for humans. To understand AsA biosynthesis in kiwifruit, AsA levels and the relative expression of genes putatively involved in AsA biosynthesis, regeneration, and transport were correlated by quantitative polymerase chain reaction in leaves and during fruit development in four kiwifruit genotypes (three species; A. eriantha, A. chinensis, and A. deliciosa). During fruit development, fruit AsA concentration peaked between 4 and 6 weeks after anthesis with A. eriantha having 3-16-fold higher AsA than other genotypes. The rise in AsA concentration typically occurred close to the peak in expression of the L-galactose pathway biosynthetic genes, particularly the GDP-L-galactose guanyltransferase gene. The high concentration of AsA found in the fruit of A. eriantha is probably due to higher expression of the GDP-mannose-3',5'-epimerase and GDP-L-galactose guanyltransferase genes. Over-expression of the kiwifruit GDP-L-galactose guanyltransferase gene in Arabidopsis resulted in up to a 4-fold increase in AsA, while up to a 7-fold increase in AsA was observed in transient expression studies where both GDP-L-galactose guanyltransferase and GDP-mannose-3',5'-epimerase genes were co-expressed. These studies show the importance of GDP-L-galactose guanyltransferase as a rate-limiting step to AsA, and demonstrate how AsA can be significantly increased in plants.
Figures

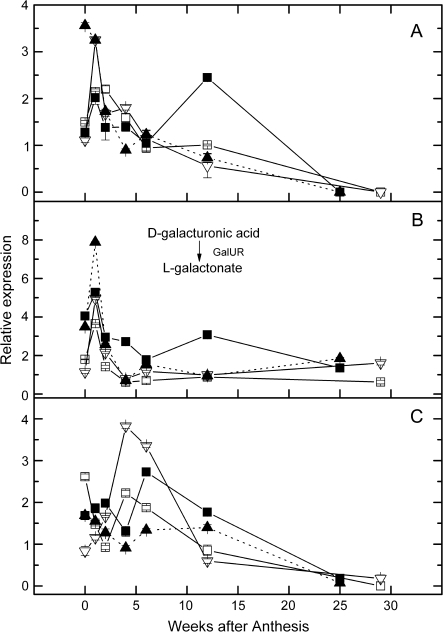
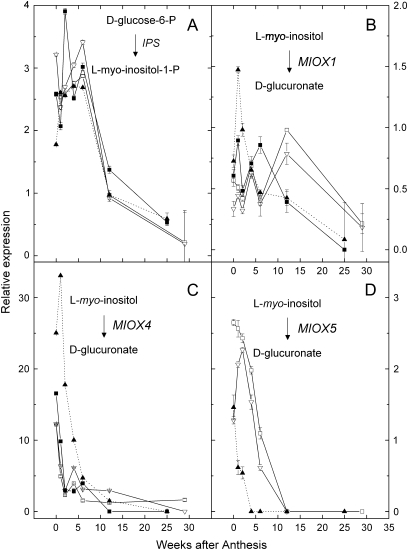
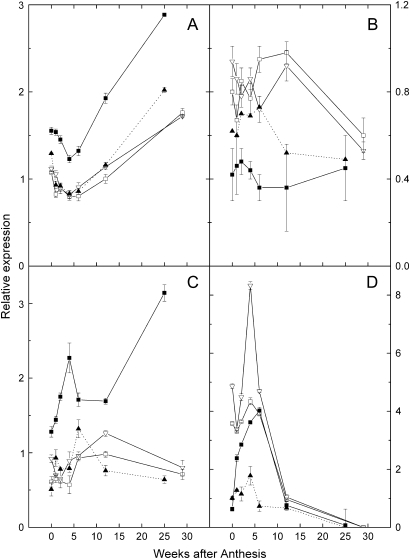
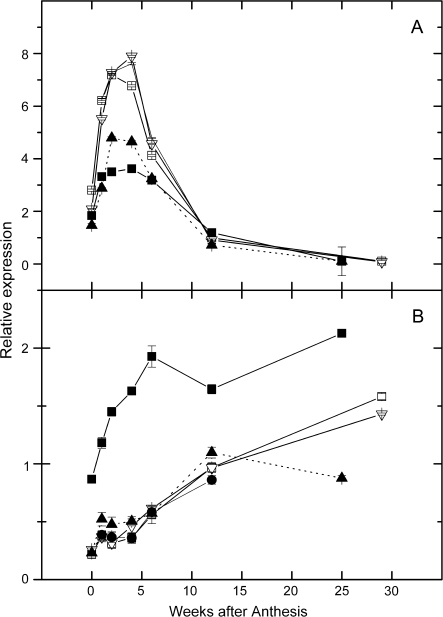
References
-
- Agius F, Gonzalez-Lamothe R, Caballero JL, Munoz-Blanco J, Botella MA, Valpuesta V. Engineering increased vitamin C levels in plants by over-expression of a D-galacturonic acid reductase. Nature Biotechnology. 2003;21:177–181. - PubMed
-
- Anderson E. The preparation of L-galactose from flax seed mucilage. Journal of Biological Chemistry. 1933;100:249–253.
-
- Arrigoni O, De Tullio MC. The role of ascorbic acid in cell metabolism: between gene-directed functions and unpredictable chemical reactions. Journal of Plant Physiology. 2000;157:481–488.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
