

Lipocalin-2 is an autocrine mediator of reactive astrocytosis
- PMID: 19129400
- PMCID: PMC6664907
- DOI: 10.1523/JNEUROSCI.5273-08.2009
Lipocalin-2 is an autocrine mediator of reactive astrocytosis
Abstract
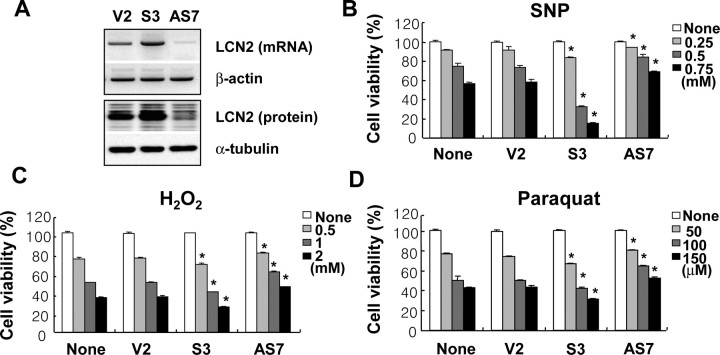
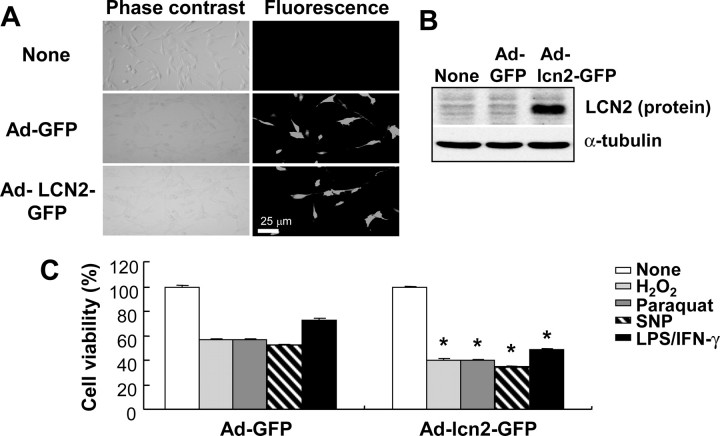
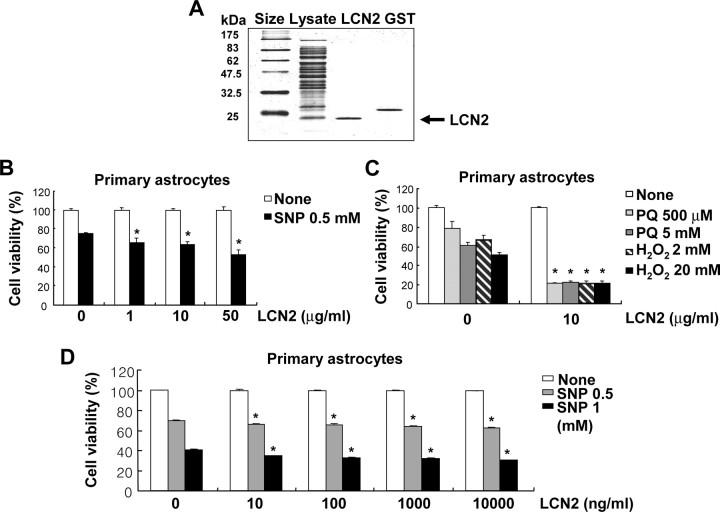
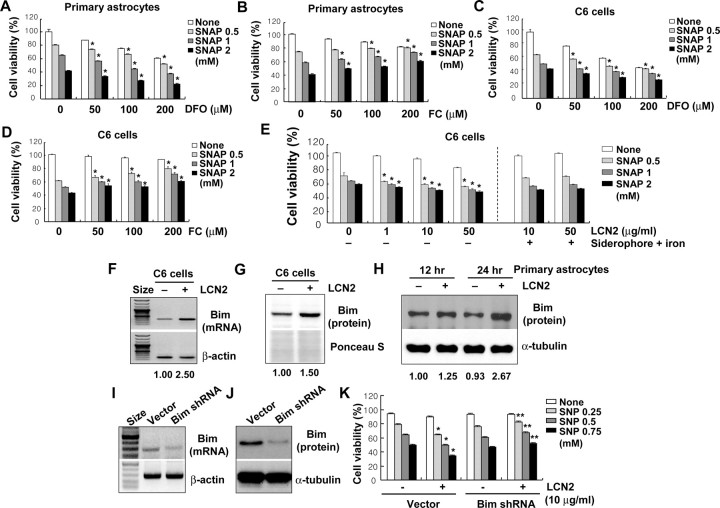
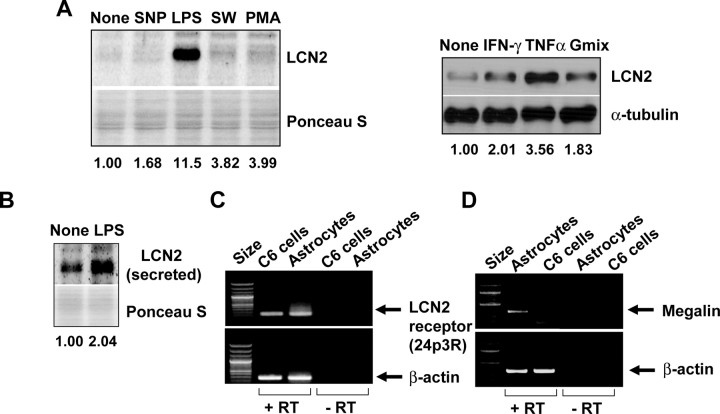
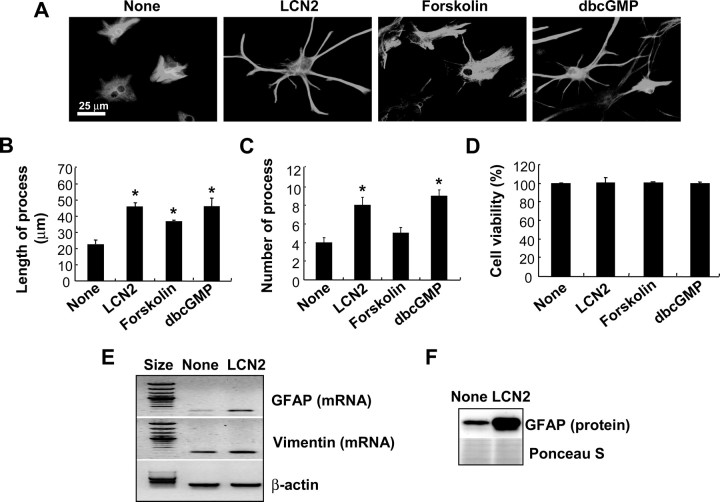
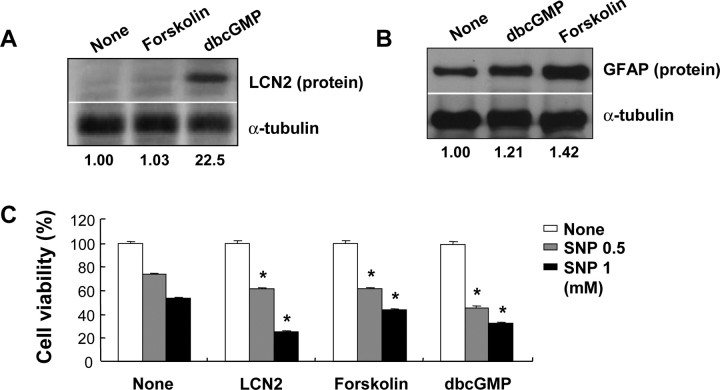
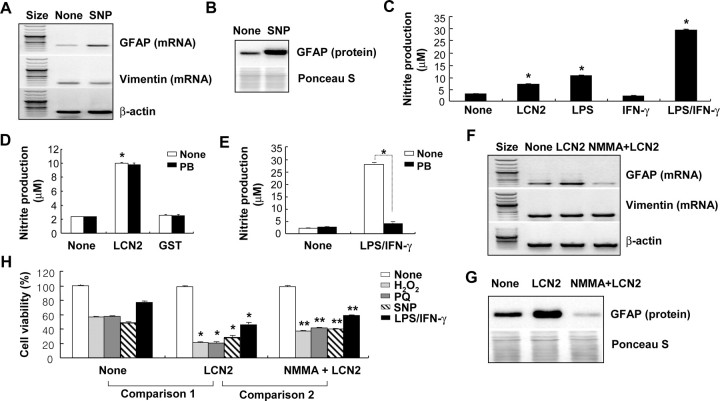
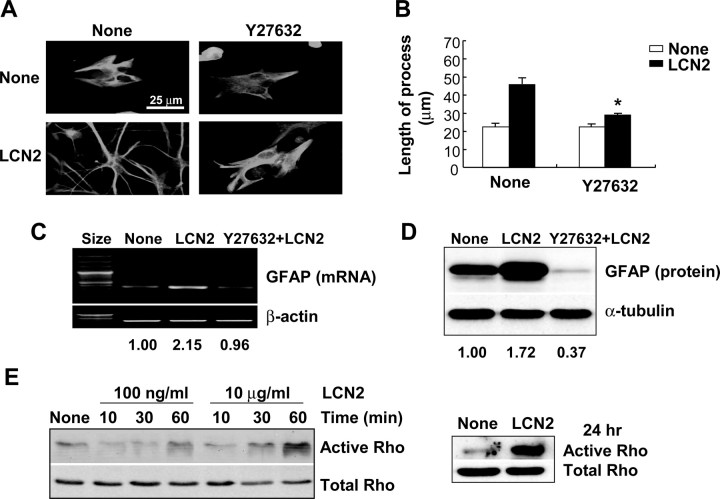
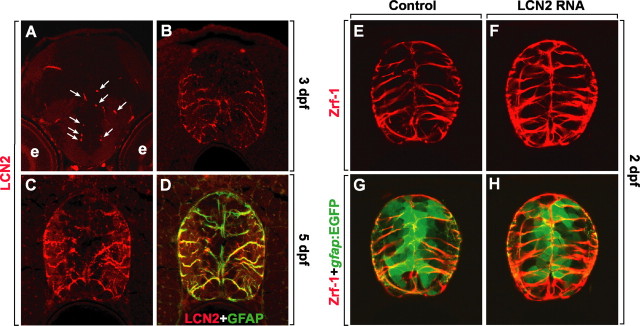
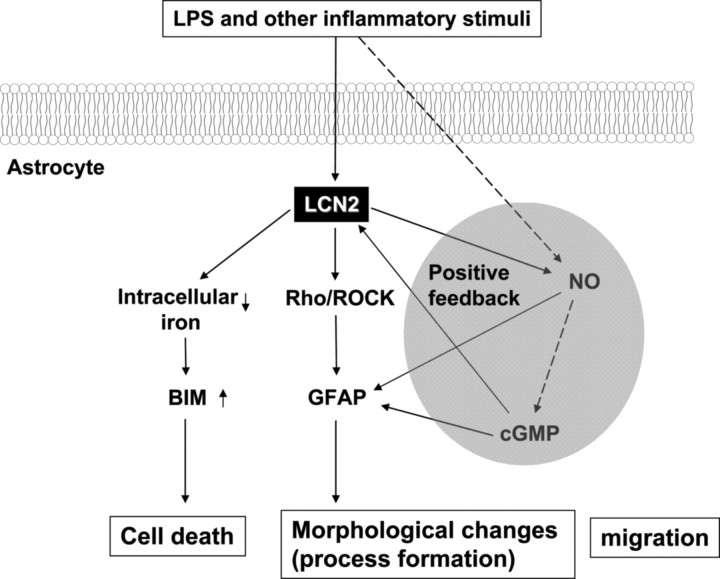
Astrocytes, the most abundant glial cell type in the brain, provide metabolic and trophic support to neurons and modulate synaptic activity. In response to a brain injury, astrocytes proliferate and become hypertrophic with an increased expression of intermediate filament proteins. This process is collectively referred to as reactive astrocytosis. Lipocalin 2 (lcn2) is a member of the lipocalin family that binds to small hydrophobic molecules. We propose that lcn2 is an autocrine mediator of reactive astrocytosis based on the multiple roles of lcn2 in the regulation of cell death, morphology, and migration of astrocytes. lcn2 expression and secretion increased after inflammatory stimulation in cultured astrocytes. Forced expression of lcn2 or treatment with LCN2 protein increased the sensitivity of astrocytes to cytotoxic stimuli. Iron and BIM (Bcl-2-interacting mediator of cell death) proteins were involved in the cytotoxic sensitization process. LCN2 protein induced upregulation of glial fibrillary acidic protein (GFAP), cell migration, and morphological changes similar to characteristic phenotypic changes termed reactive astrocytosis. The lcn2-induced phenotypic changes of astrocytes occurred through a Rho-ROCK (Rho kinase)-GFAP pathway, which was positively regulated by nitric oxide and cGMP. In zebrafishes, forced expression of rat lcn2 gene increased the number and thickness of cellular processes in GFAP-expressing radial glia cells, suggesting that lcn2 expression in glia cells plays an important role in vivo. Our results suggest that lcn2 acts in an autocrine manner to induce cell death sensitization and morphological changes in astrocytes under inflammatory conditions and that these phenotypic changes may be the basis of reactive astrocytosis in vivo.
Figures

References
-
- Abe K, Misawa M. Astrocyte stellation induced by Rho kinase inhibitors in culture. Brain Res Dev Brain Res. 2003;143:99–104. - PubMed
-
- Anzenberger U, Bit-Avragim N, Rohr S, Rudolph F, Dehmel B, Willnow TE, Abdelilah-Seyfried S. Elucidation of megalin/LRP2-dependent endocytic transport processes in the larval zebrafish pronephros. J Cell Sci. 2006;119:2127–2137. - PubMed
-
- Aschner M. Astrocytes as mediators of immune and inflammatory responses in the CNS. Neurotoxicology. 1998;19:269–281. - PubMed
-
- Avalos AM, Arthur WT, Schneider P, Quest AF, Burridge K, Leyton L. Aggregation of integrins and RhoA activation are required for Thy-1-induced morphological changes in astrocytes. J Biol Chem. 2004;279:39139–39145. - PubMed
-
- Barres BA, Barde Y. Neuronal and glial cell biology. Curr Opin Neurobiol. 2000;10:642–648. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous