

Intersubunit coordination in a homomeric ring ATPase
- PMID: 19129763
- PMCID: PMC2716090
- DOI: 10.1038/nature07637
Intersubunit coordination in a homomeric ring ATPase
Abstract
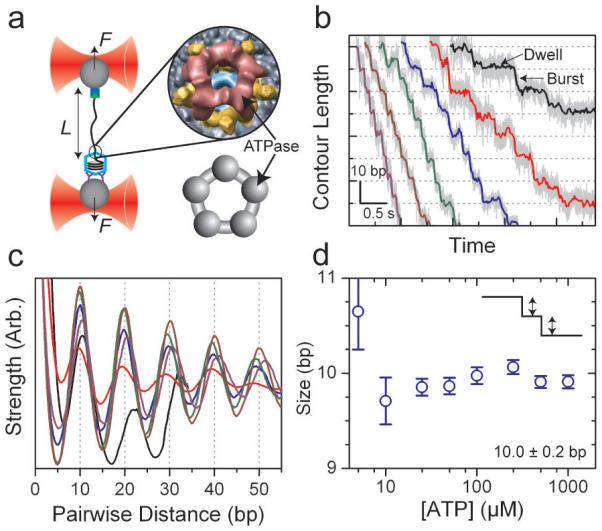
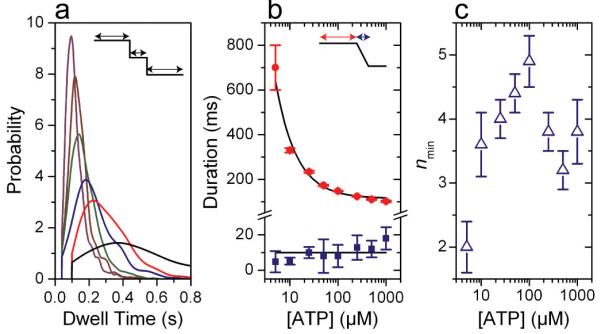
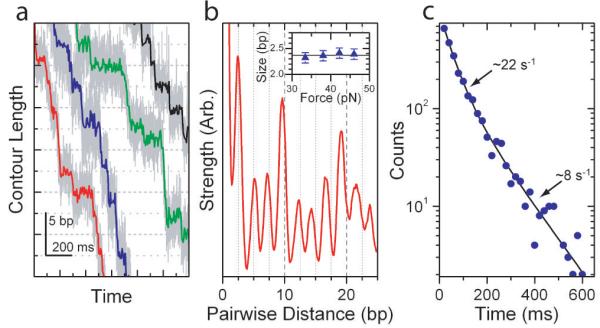
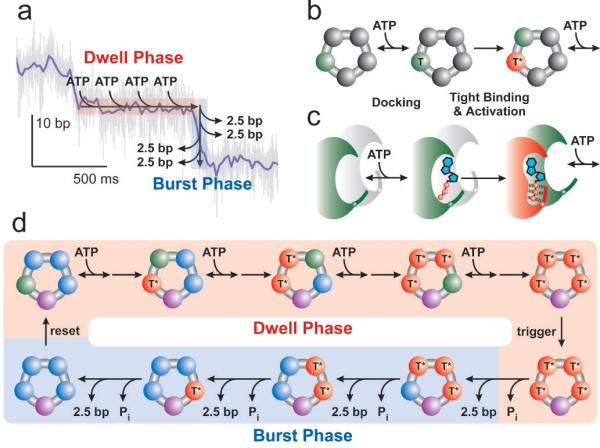
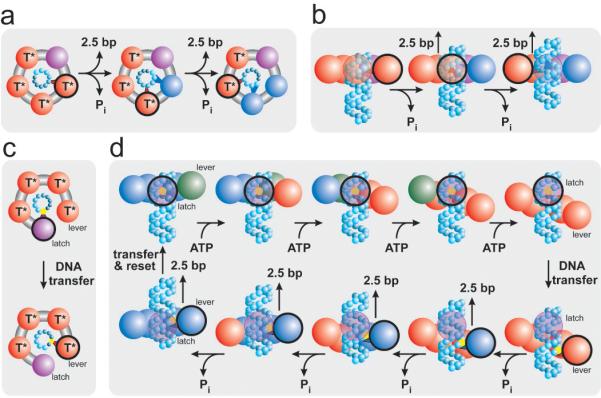
Homomeric ring ATPases perform many vital and varied tasks in the cell, ranging from chromosome segregation to protein degradation. Here we report the direct observation of the intersubunit coordination and step size of such a ring ATPase, the double-stranded-DNA packaging motor in the bacteriophage phi29. Using high-resolution optical tweezers, we find that packaging occurs in increments of 10 base pairs (bp). Statistical analysis of the preceding dwell times reveals that multiple ATPs bind during each dwell, and application of high force reveals that these 10-bp increments are composed of four 2.5-bp steps. These results indicate that the hydrolysis cycles of the individual subunits are highly coordinated by means of a mechanism novel for ring ATPases. Furthermore, a step size that is a non-integer number of base pairs demands new models for motor-DNA interactions.
Figures
Comment in
-
Molecular biology: Concealed enzyme coordination.Nature. 2009 Jan 22;457(7228):392-3. doi: 10.1038/457392a. Nature. 2009. PMID: 19158782 No abstract available.
References
-
- Latterich M, Patel S. The AAA team: related ATPases with diverse functions. Trends in Cell Biology. 1998;8:65. - PubMed
-
- Ogura T, Wilkinson AJ. AAA+ superfamily ATPases: common structure--diverse function. Genes to Cells. 2001;6:575–597. - PubMed
-
- Iyer LM, Leipe DD, Koonin EV, Aravind L. Evolutionary history and higher order classification of AAA+ ATPases. J Struct Biol. 2004;146:11–31. - PubMed
-
- Erzberger JP, Berger JM. Evolutionary relationships and structural mechanisms of AAA+ proteins. Annu Rev Biophys Biomol Struct. 2006;35:93–114. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
