

Drosophila PTB promotes formation of high-order RNP particles and represses oskar translation
- PMID: 19131435
- PMCID: PMC2648539
- DOI: 10.1101/gad.505709
Drosophila PTB promotes formation of high-order RNP particles and represses oskar translation
Abstract
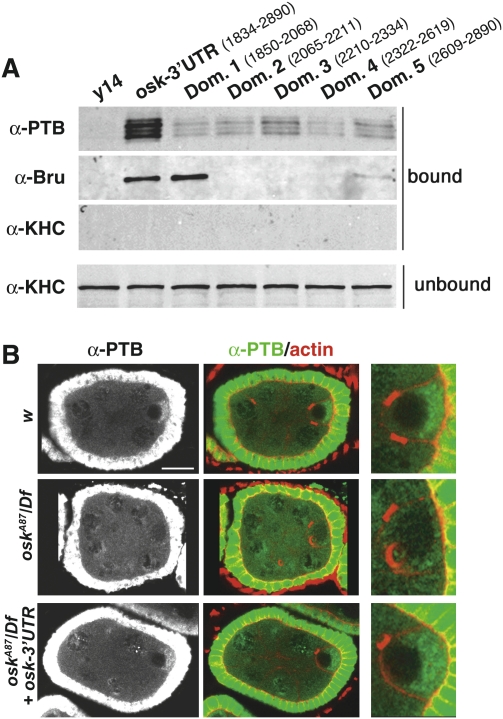
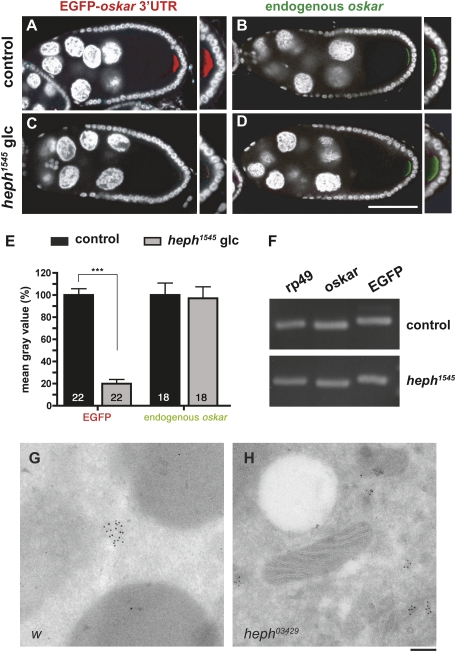
Local translation of asymmetrically enriched mRNAs is a powerful mechanism for functional polarization of the cell. In Drosophila, exclusive accumulation of Oskar protein at the posterior pole of the oocyte is essential for development of the future embryo. This is achieved by the formation of a dynamic oskar ribonucleoprotein (RNP) complex regulating the transport of oskar mRNA, its translational repression while unlocalized, and its translational activation upon arrival at the posterior pole. We identified the nucleo-cytoplasmic shuttling protein PTB (polypyrimidine tract-binding protein)/hnRNP I as a new factor associating with the oskar RNP in vivo. While PTB function is largely dispensable for oskar mRNA transport, it is necessary for translational repression of the localizing mRNA. Unexpectedly, a cytoplasmic form of PTB can associate with oskar mRNA and repress its translation, suggesting that nuclear recruitment of PTB to oskar complexes is not required for its regulatory function. Furthermore, PTB binds directly to multiple sites along the oskar 3' untranslated region and mediates assembly of high-order complexes containing multiple oskar RNA molecules in vivo. Thus, PTB is a key structural component of oskar RNP complexes that dually controls formation of high-order RNP particles and translational silencing.
Figures

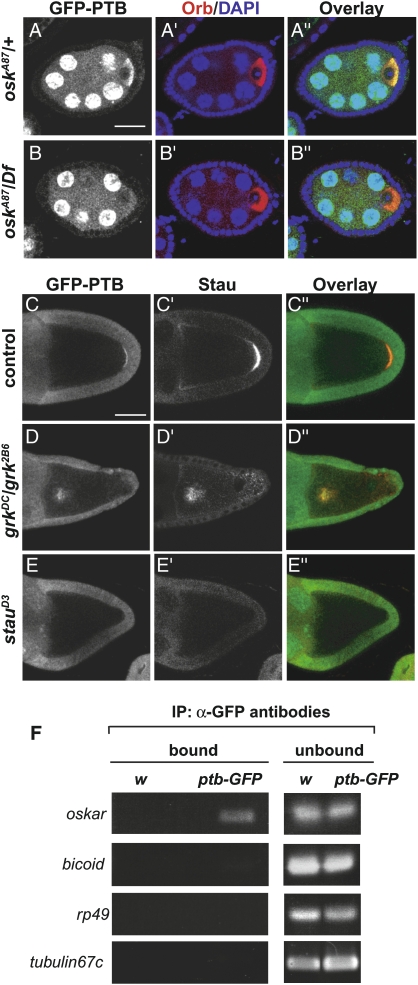
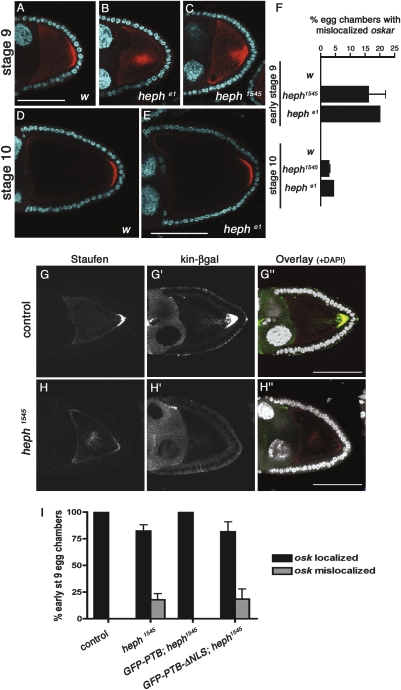
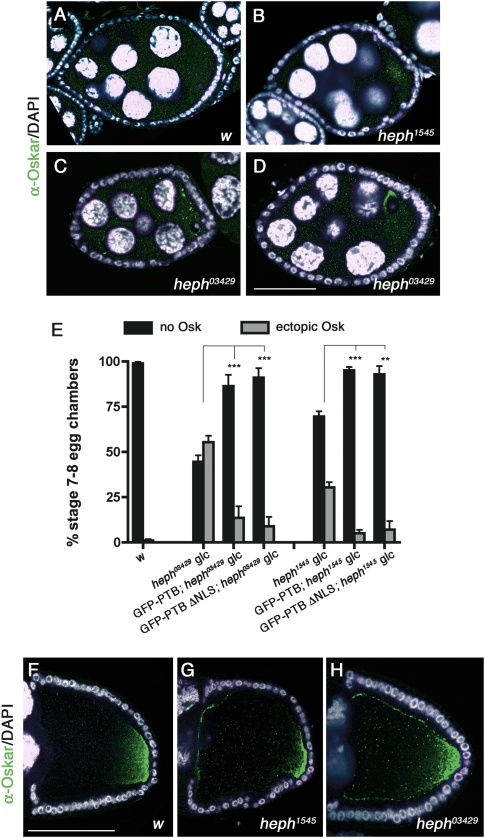
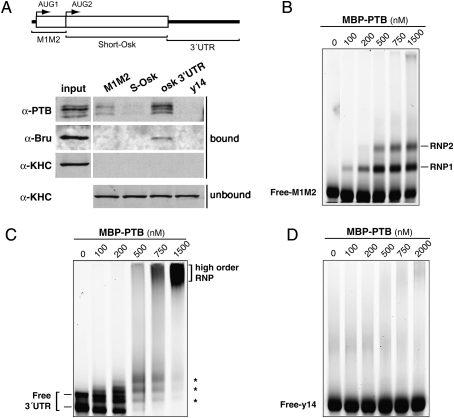
Comment in
-
A splicer that represses (translation).Genes Dev. 2009 Jan 15;23(2):133-7. doi: 10.1101/gad.1768509. Genes Dev. 2009. PMID: 19171776
References
-
- Chang J.S., Tan L., Schedl P. The Drosophila CPEB homolog, Orb, is required for Oskar protein expression in oocytes. Dev. Biol. 1999;215:91–106. - PubMed
-
- Chekulaeva M., Hentze M.W., Ephrussi A. Bruno acts as a dual repressor of oskar translation, promoting mRNA oligomerization and formation of silencing particles. Cell. 2006;124:521–533. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases