

Immunobiology of human cytomegalovirus: from bench to bedside
- PMID: 19136435
- PMCID: PMC2620639
- DOI: 10.1128/CMR.00034-08
Immunobiology of human cytomegalovirus: from bench to bedside
Abstract
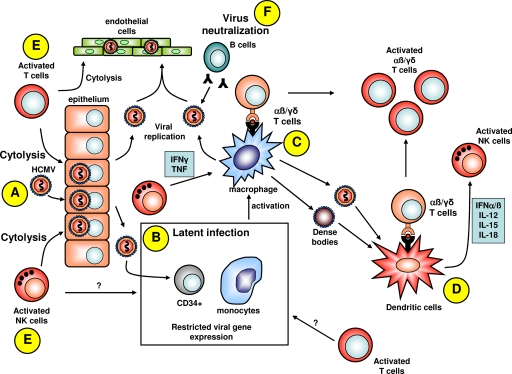
Following primary infection, human cytomegalovirus (HCMV) establishes lifelong latency and periodically reactivates without causing symptoms in healthy individuals. In the absence of an adequate host-derived immune response, this fine balance of permitting viral reactivation without causing pathogenesis is disrupted, and HCMV can subsequently cause invasive disease and an array of damaging indirect immunological effects. Over the last decade, our knowledge of the immune response to HCMV infection in healthy virus carriers and diseased individuals has allowed us to translate these findings to develop better diagnostic tools and therapeutic strategies. The application of these emerging technologies in the clinical setting is likely to provide opportunities for better management of patients with HCMV-associated diseases.
Figures
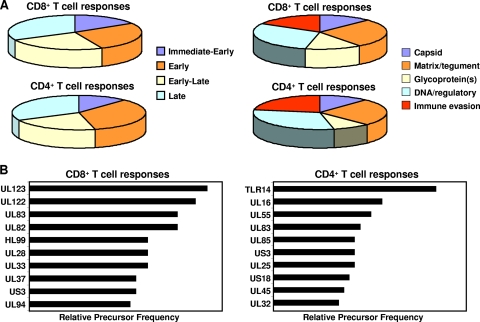
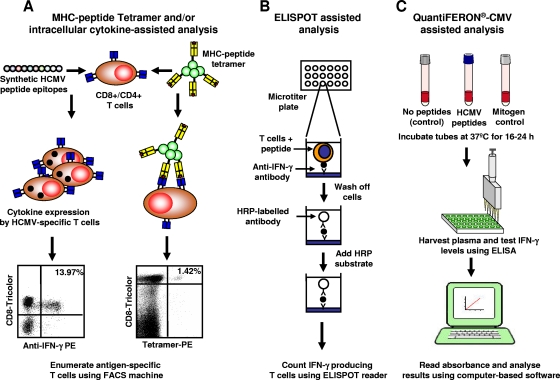
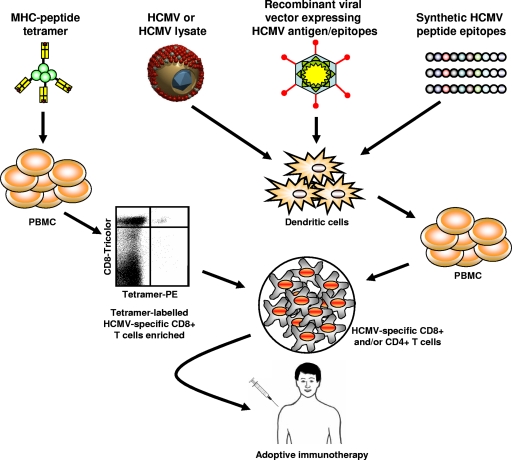
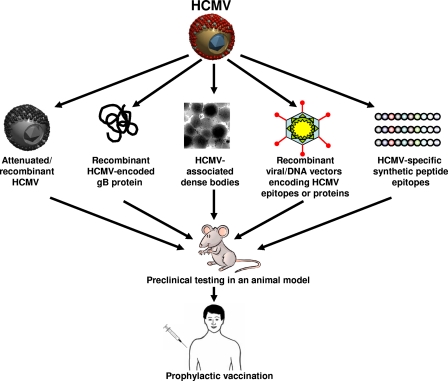
References
-
- Adler, S. P., S. H. Hempfling, S. E. Starr, S. A. Plotkin, and S. Riddell. 1998. Safety and immunogenicity of the Towne strain cytomegalovirus vaccine. Pediatr. Infect. Dis. J. 17:200-206. - PubMed
-
- Adler, S. P., G. Nigro, and L. Pereira. 2007. Recent advances in the prevention and treatment of congenital cytomegalovirus infections. Semin. Perinatol. 31:10-18. - PubMed
-
- Adler, S. P., S. A. Plotkin, E. Gonczol, M. Cadoz, C. Meric, J. B. Wang, P. Dellamonica, A. M. Best, J. Zahradnik, S. Pincus, K. Berencsi, W. I. Cox, and Z. Gyulai. 1999. A canarypox vector expressing cytomegalovirus (CMV) glycoprotein B primes for antibody responses to a live attenuated CMV vaccine (Towne). J. Infect. Dis. 180:843-846. - PubMed
-
- Ahn, K., A. Gruhler, B. Galocha, T. R. Jones, E. J. Wiertz, H. L. Ploegh, P. A. Peterson, Y. Yang, and K. Fruh. 1997. The ER-luminal domain of the HCMV glycoprotein US6 inhibits peptide translocation by TAP. Immunity 6:613-621. - PubMed
-
- Akbar, A. N., and J. M. Fletcher. 2005. Memory T cell homeostasis and senescence during aging. Curr. Opin. Immunol. 17:480-485. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
