

Modulation of gene expression in U251 glioblastoma cells by binding of mutant p53 R273H to intronic and intergenic sequences
- PMID: 19139068
- PMCID: PMC2655687
- DOI: 10.1093/nar/gkn1085
Modulation of gene expression in U251 glioblastoma cells by binding of mutant p53 R273H to intronic and intergenic sequences
Abstract
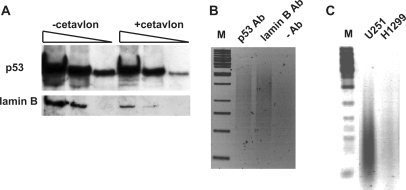
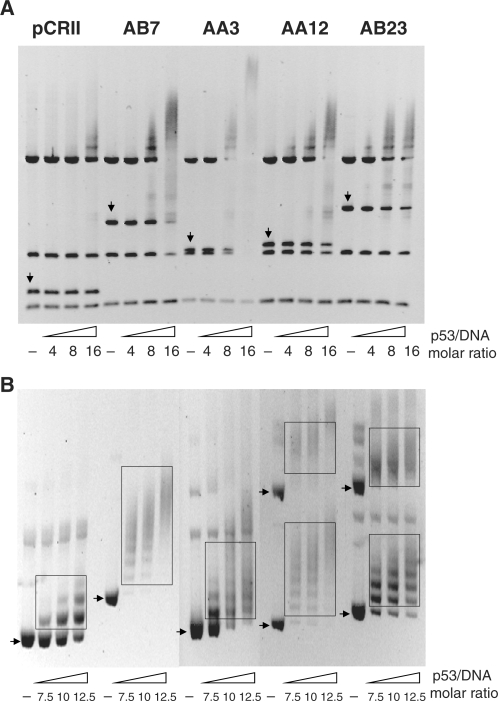
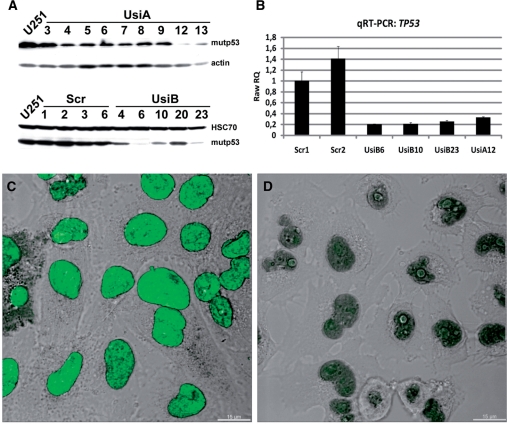
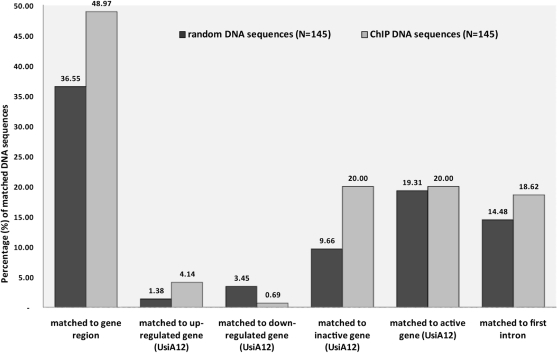
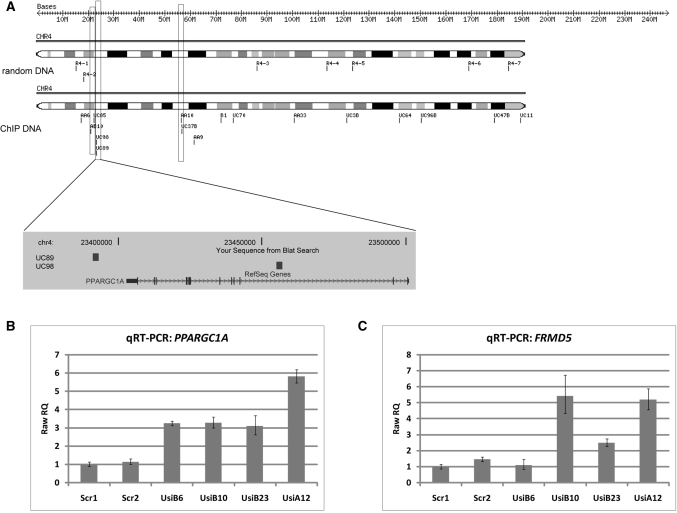
Missense point mutations in the TP53 gene are frequent genetic alterations in human tumor tissue and cell lines derived thereof. Mutant p53 (mutp53) proteins have lost sequence-specific DNA binding, but have retained the ability to interact in a structure-selective manner with non-B DNA and to act as regulators of transcription. To identify functional binding sites of mutp53, we established a small library of genomic sequences bound by p53(R273H) in U251 human glioblastoma cells using chromatin immunoprecipitation (ChIP). Mutp53 binding to isolated DNA fragments confirmed the specificity of the ChIP. The mutp53 bound DNA sequences are rich in repetitive DNA elements, which are dispersed over non-coding DNA regions. Stable down-regulation of mutp53 expression strongly suggested that mutp53 binding to genomic DNA is functional. We identified the PPARGC1A and FRMD5 genes as p53(R273H) targets regulated by binding to intronic and intra-genic sequences. We propose a model that attributes the oncogenic functions of mutp53 to its ability to interact with intronic and intergenic non-B DNA sequences and modulate gene transcription via re-organization of chromatin.
Figures


References
-
- Soussi T, Kato S, Levy PP, Ishioka C. Reassessment of the TP53 mutation database in human disease by data mining with a library of TP53 missense mutations. Hum. Mutat. 2005;25:6–17. - PubMed
-
- Petitjean A, Mathe E, Kato S, Ishioka C, Tavtigian SV, Hainaut P, Olivier M. Impact of mutant p53 functional properties on TP53 mutation patterns and tumor phenotype: lessons from recent developments in the IARC TP53 database. Hum. Mutat. 2007;28:622–629. - PubMed
-
- Petitjean A, Achatz MI, Borresen-Dale AL, Hainaut P, Olivier M. TP53 mutations in human cancers: functional selection and impact on cancer prognosis and outcomes. Oncogene. 2007;26:2157–2165. - PubMed
-
- Ang HC, Joerger AC, Mayer S, Fersht AR. Effects of common cancer mutations on stability and DNA binding of full-length p53 compared with isolated core domains. J. Biol. Chem. 2006;281:21934–21941. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
