

Combinatorial assembly of simple and complex D-lysergic acid alkaloid peptide classes in the ergot fungus Claviceps purpurea
- PMID: 19139103
- PMCID: PMC2652330
- DOI: 10.1074/jbc.M807168200
Combinatorial assembly of simple and complex D-lysergic acid alkaloid peptide classes in the ergot fungus Claviceps purpurea
Abstract
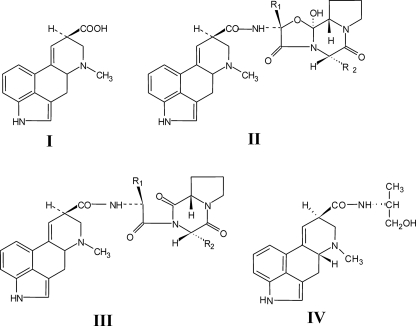
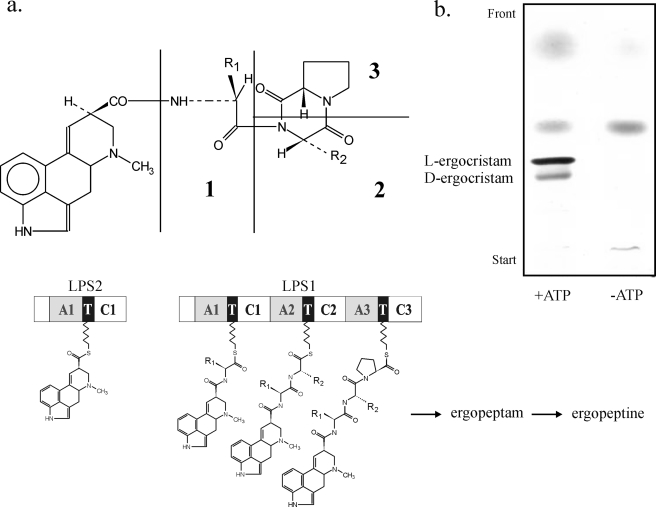
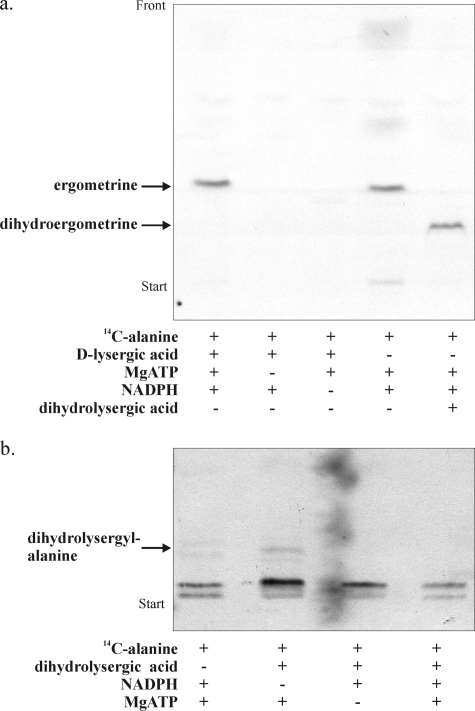
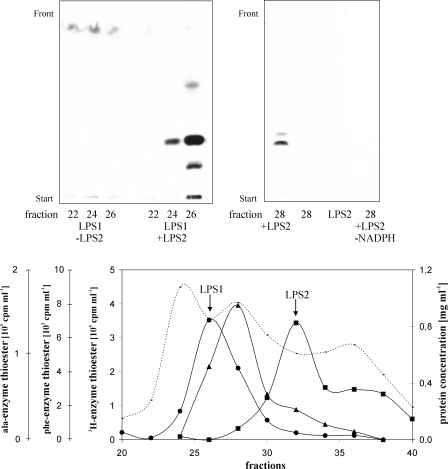
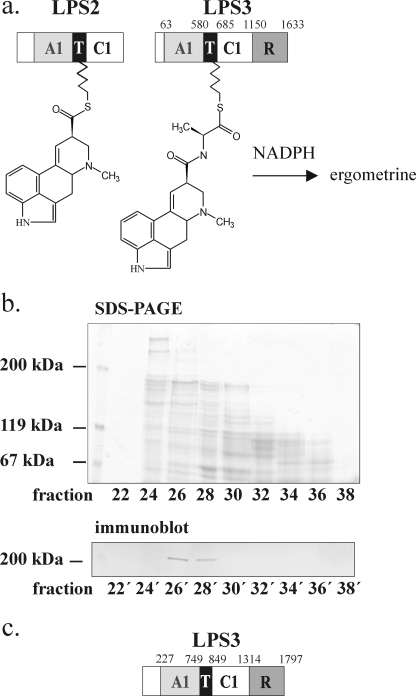
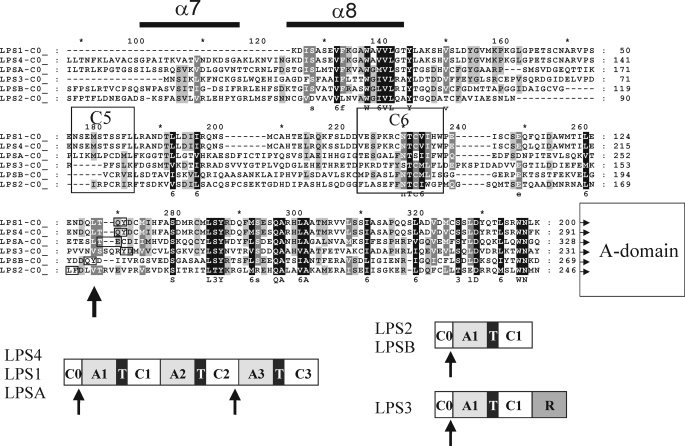
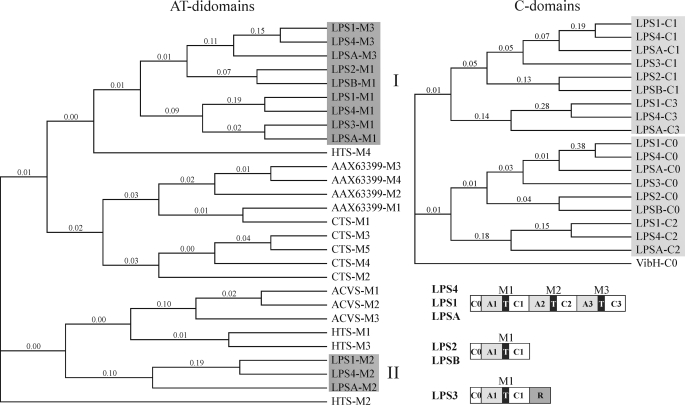
The ergot fungus Claviceps purpurea produces both ergopeptines and simple d-lysergic acid alkylamides. In the ergopeptines, such as ergotamine, d-lysergic acid is linked to a bicyclic tripeptide in amide-like fashion, whereas in the d-lysergylalkanolamides it is linked to an amino alcohol derived from alanine. We show here that these compound classes are synthesized by a set of three non-ribosomal lysergyl peptide synthetases (LPSs), which interact in a combinatorial fashion for synthesis of the relevant product. The trimodular LPS1 assembles with LPS2, the d-lysergic acid recruiting module, to synthesize the d-lysergyltripeptide precursors of ergopeptines from d-lysergic acid and the three amino acids of the peptide chain. Alternatively, LPS2 can assemble with a distinct monomodular non-ribosomal peptide synthetase (NRPS) subunit (ergometrine synthetase) to synthesize the d-lysergic acid alkanolamide ergometrine from d-lysergic acid and alanine. The synthesis proceeds via covalently bound d-lysergyl alanine and release of dipeptide as alcohol with consumption of NADPH. Enzymatic and immunochemical analyses showed that ergometrine synthetase is most probably the enzyme LPS3 whose gene had been identified previously as part of the ergot alkaloid biosynthesis gene cluster in C. purpurea. Inspections of all LPS sequences showed no recognizable peptide linkers for their protein-protein interactions as in NRPS subunits of bacteria. Instead, they all carry conserved N-terminal domains (C0-domains) with similarity to the C-terminal halves of NRPS condensation domains pointing to an alternative mechanism of subunit-subunit interactions in fungal NRPS systems. Phylogenetic analysis of LPS modules and the C0-domains suggests that these enzyme systems most probably evolved by module duplications and rearrangements from a bimodular ancestor.
Figures
Similar articles
-
Molecular cloning and analysis of the ergopeptine assembly system in the ergot fungus Claviceps purpurea.Chem Biol. 2003 Dec;10(12):1281-92. doi: 10.1016/j.chembiol.2003.11.013. Chem Biol. 2003. PMID: 14700635
-
Mechanism of alkaloid cyclopeptide synthesis in the ergot fungus Claviceps purpurea.Chem Biol. 1997 Mar;4(3):223-30. doi: 10.1016/s1074-5521(97)90292-1. Chem Biol. 1997. PMID: 9115414
-
D-Lysergyl peptide synthetase from the ergot fungus Claviceps purpurea.J Biol Chem. 1996 Nov 1;271(44):27524-30. doi: 10.1074/jbc.271.44.27524. J Biol Chem. 1996. PMID: 8910337
-
Ergot alkaloids--biology and molecular biology.Alkaloids Chem Biol. 2006;63:45-86. doi: 10.1016/s1099-4831(06)63002-2. Alkaloids Chem Biol. 2006. PMID: 17133714 Review.
-
Methods of Lysergic Acid Synthesis-The Key Ergot Alkaloid.Molecules. 2022 Oct 28;27(21):7322. doi: 10.3390/molecules27217322. Molecules. 2022. PMID: 36364148 Free PMC article. Review.
Cited by
-
Branching and converging pathways in fungal natural product biosynthesis.Fungal Biol Biotechnol. 2022 Mar 7;9(1):6. doi: 10.1186/s40694-022-00135-w. Fungal Biol Biotechnol. 2022. PMID: 35255990 Free PMC article. Review.
-
A Baeyer-Villiger Monooxygenase Gene Involved in the Synthesis of Lysergic Acid Amides Affects the Interaction of the Fungus Metarhizium brunneum with Insects.Appl Environ Microbiol. 2021 Aug 11;87(17):e0074821. doi: 10.1128/AEM.00748-21. Epub 2021 Aug 11. Appl Environ Microbiol. 2021. PMID: 34160271 Free PMC article.
-
Independent Evolution of a Lysergic Acid Amide in Aspergillus Species.Appl Environ Microbiol. 2021 Nov 24;87(24):e0180121. doi: 10.1128/AEM.01801-21. Epub 2021 Sep 29. Appl Environ Microbiol. 2021. PMID: 34586904 Free PMC article.
-
Transcription factor Xpp1 is a switch between primary and secondary fungal metabolism.Proc Natl Acad Sci U S A. 2017 Jan 24;114(4):E560-E569. doi: 10.1073/pnas.1609348114. Epub 2017 Jan 10. Proc Natl Acad Sci U S A. 2017. PMID: 28074041 Free PMC article.
-
Unveiling nonribosomal peptide synthetases from the ergot fungus Claviceps purpurea involved in the formation of diverse ergopeptines.Acta Pharm Sin B. 2025 Jun;15(6):3321-3337. doi: 10.1016/j.apsb.2025.03.022. Epub 2025 Mar 13. Acta Pharm Sin B. 2025. PMID: 40654343 Free PMC article.
References
-
- Stadler, P. A., and Giger, R. (1984) in Natural Products and Drug Development (Krosgard-Larson, P., and Kofod, H., eds) pp. 463–485, Munksgaard, Copenhagen, Denmark
-
- Burkhalter, A., Julius, D. J., and Katzung, B. G. (1998) in Basic and Clinical Pharmacology (Katzung, B. G., ed) pp. 261–286, Appleton-Lange, New York
-
- Berde, B., and Stuermer, E. (1978) in Introduction to the Pharmacology of Ergot Alkaloids and Related Compounds (Aellig, W. H., Berde, B., and Schild, O. H., eds) pp. 1–28, Springer, Berlin
-
- Flieger, M., Wurst, M., and Shelby, R. (1997) Folia Microbiol. 42 3–29 - PubMed
-
- Kobel, H., and Sanglier, J. J. (1986) in Biotechnology (Rehm, H. J., and Reeds, G., eds) pp. 569–609, VCH Verlagsgesellschaft, Weinheim, Germany
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
