

Estrogen directly activates AID transcription and function
- PMID: 19139166
- PMCID: PMC2626679
- DOI: 10.1084/jem.20080521
Estrogen directly activates AID transcription and function
Abstract
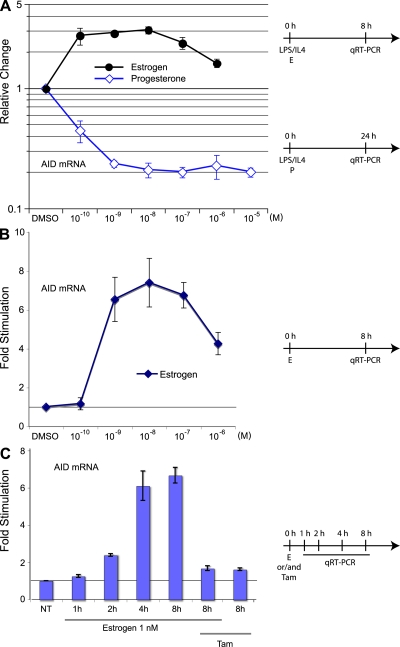
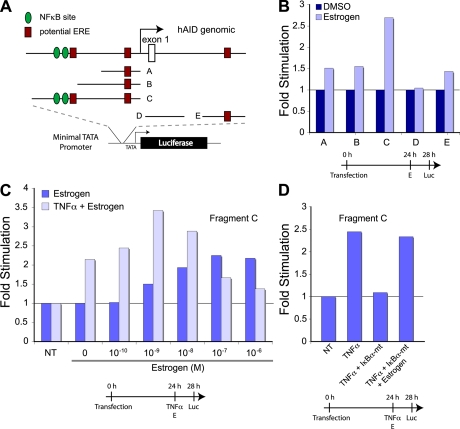
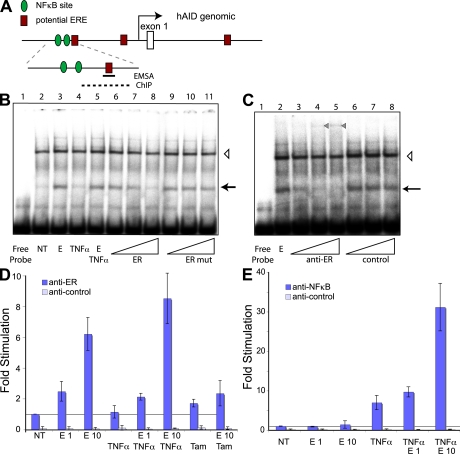
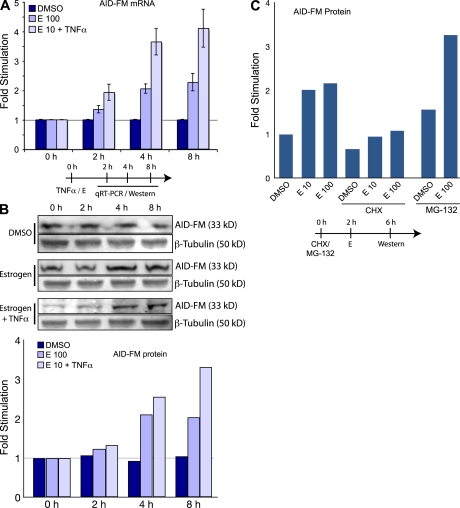
The immunological targets of estrogen at the molecular, humoral, and cellular level have been well documented, as has estrogen's role in establishing a gender bias in autoimmunity and cancer. During a healthy immune response, activation-induced deaminase (AID) deaminates cytosines at immunoglobulin (Ig) loci, initiating somatic hypermutation (SHM) and class switch recombination (CSR). Protein levels of nuclear AID are tightly controlled, as unregulated expression can lead to alterations in the immune response. Furthermore, hyperactivation of AID outside the immune system leads to oncogenesis. Here, we demonstrate that the estrogen-estrogen receptor complex binds to the AID promoter, enhancing AID messenger RNA expression, leading to a direct increase in AID protein production and alterations in SHM and CSR at the Ig locus. Enhanced translocations of the c-myc oncogene showed that the genotoxicity of estrogen via AID production was not limited to the Ig locus. Outside of the immune system (e.g., breast and ovaries), estrogen induced AID expression by >20-fold. The estrogen response was also partially conserved within the DNA deaminase family (APOBEC3B, -3F, and -3G), and could be inhibited by tamoxifen, an estrogen antagonist. We therefore suggest that estrogen-induced autoimmunity and oncogenesis may be derived through AID-dependent DNA instability.
Figures
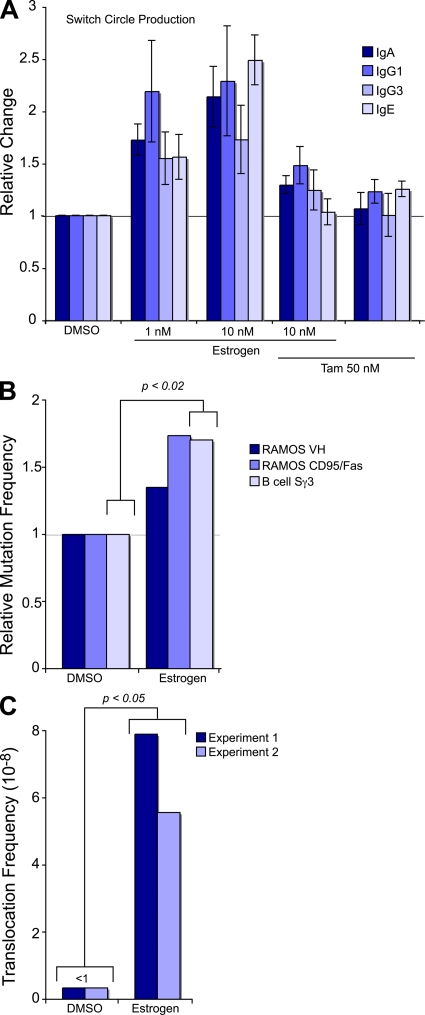
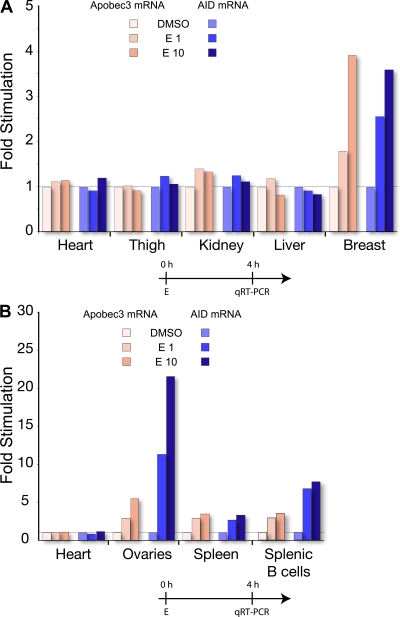
Comment in
-
Women, autoimmunity, and cancer: a dangerous liaison between estrogen and activation-induced deaminase?J Exp Med. 2009 Jan 16;206(1):11-3. doi: 10.1084/jem.20080086. Epub 2009 Jan 12. J Exp Med. 2009. PMID: 19139165 Free PMC article.
References
-
- Neuberger, M.S., R.S. Harris, J. Di Noia, and S.K. Petersen-Mahrt. 2003. Immunity through DNA deamination. Trends Biochem. Sci. 28:305–312. - PubMed
-
- Arakawa, H., J. Hauschild, and J.M. Buerstedde. 2002. Requirement of the activation-induced deaminase (AID) gene for immunoglobulin gene conversion. Science. 295:1301–1306. - PubMed
-
- Harris, R.S., J.E. Sale, S.K. Petersen-Mahrt, and M.S. Neuberger. 2002. AID is essential for immunoglobulin V gene conversion in a cultured B cell line. Curr. Biol. 12:435–438. - PubMed
-
- Muramatsu, M., K. Kinoshita, S. Fagarasan, S. Yamada, Y. Shinkai, and T. Honjo. 2000. Class switch recombination and hypermutation require activation-induced cytidine deaminase (AID), a potential RNA editing enzyme. Cell. 102:553–563. - PubMed
-
- Muramatsu, M., V.S. Sankaranand, S. Anant, M. Sugai, K. Kinoshita, N.O. Davidson, and T. Honjo. 1999. Specific expression of activation-induced cytidine deaminase (AID), a novel member of the RNA-editing deaminase family in germinal center B cells. J. Biol. Chem. 274:18470–18476. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
