

The Epstein-Barr virus nuclear antigen-1 promotes genomic instability via induction of reactive oxygen species
- PMID: 19139406
- PMCID: PMC2650153
- DOI: 10.1073/pnas.0810619106
The Epstein-Barr virus nuclear antigen-1 promotes genomic instability via induction of reactive oxygen species
Abstract
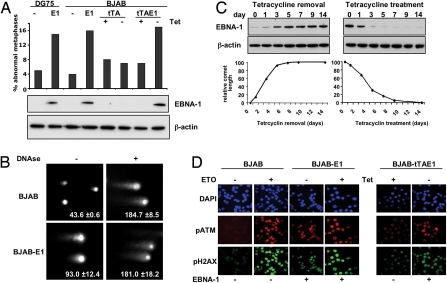
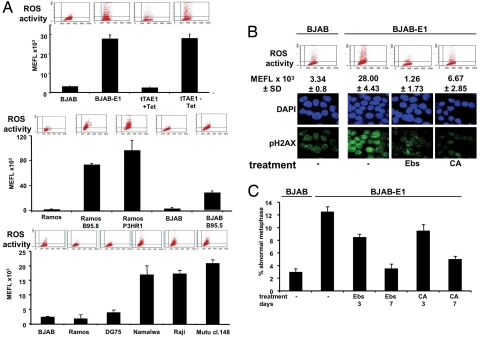
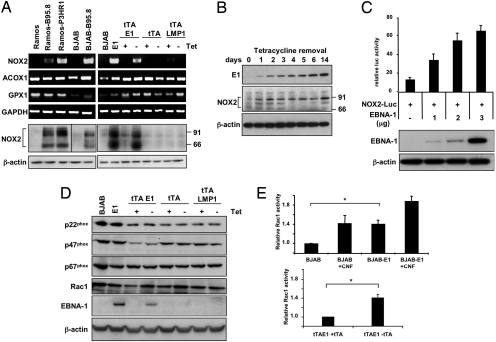
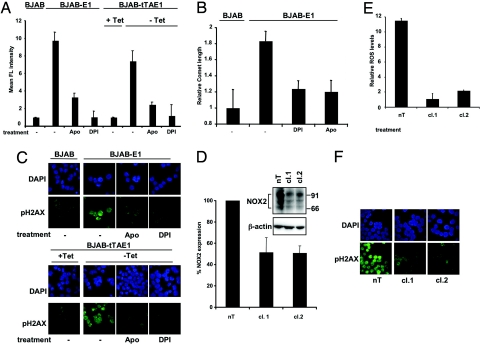
The Epstein-Barr virus (EBV) nuclear antigen (EBNA)-1 is the only viral protein expressed in all EBV-carrying malignancies, but its contribution to oncogenesis has remained enigmatic. We show that EBNA-1 induces chromosomal aberrations, DNA double-strand breaks, and engagement of the DNA damage response (DDR). These signs of genomic instability are associated with the production of reactive oxygen species (ROS) and are reversed by antioxidants. The catalytic subunit of the leukocyte NADPH oxidase, NOX2/gp91(phox), is transcriptionally activated in EBNA-1-expressing cells, whereas inactivation of the enzyme by chemical inhibitors or RNAi halts ROS production and DDR. These findings highlight a novel function of EBNA-1 and a possible mechanism by which expression of this viral protein could contribute to malignant transformation and tumor progression.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Is the Epstein-Barr virus EBNA-1 protein an oncogen?Proc Natl Acad Sci U S A. 2009 Feb 17;106(7):2091-2. doi: 10.1073/pnas.0812575106. Epub 2009 Feb 11. Proc Natl Acad Sci U S A. 2009. PMID: 19211802 Free PMC article. No abstract available.
References
-
- Kieff E, Liebowitz D. In: Virology. 2nd Ed. Fields B, Knipe D, editors. New York: Raven Press; 1990. pp. 1889–1920.
-
- Young LS, Rickinson AB. Epstein–Barr virus: 40 years on. Nat Rev Cancer. 2004;4:757–768. - PubMed
-
- Leight ER, Sugden B. EBNA-1: A protein pivotal to latent infection by Epstein–Barr virus. Rev Med Virol. 2000;10:83–100. - PubMed
-
- Hong M, et al. Suppression of Epstein–Barr nuclear antigen 1 (EBNA1) by RNA interference inhibits proliferation of EBV-positive Burkitt's lymphoma cells. J Cancer Res Clin Oncol. 2006;132:1–8. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
