

Hitch-hiking parasitic wasp learns to exploit butterfly antiaphrodisiac
- PMID: 19139416
- PMCID: PMC2630094
- DOI: 10.1073/pnas.0812277106
Hitch-hiking parasitic wasp learns to exploit butterfly antiaphrodisiac
Abstract
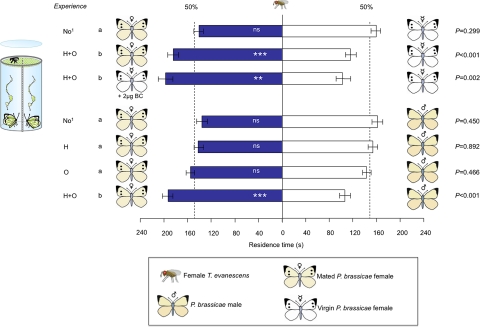
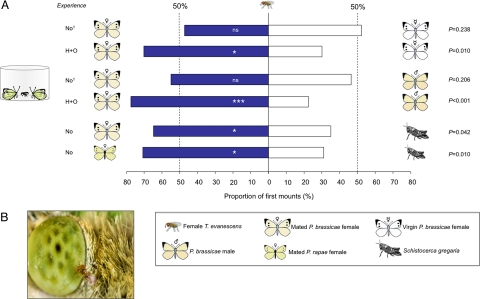
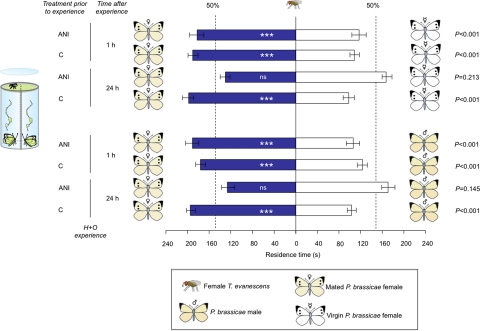
Many insects possess a sexual communication system that is vulnerable to chemical espionage by parasitic wasps. We recently discovered that a hitch-hiking (H) egg parasitoid exploits the antiaphrodisiac pheromone benzyl cyanide (BC) of the Large Cabbage White butterfly Pieris brassicae. This pheromone is passed from male butterflies to females during mating to render them less attractive to conspecific males. When the tiny parasitic wasp Trichogramma brassicae detects the antiaphrodisiac, it rides on a mated female butterfly to a host plant and then parasitizes her freshly laid eggs. The present study demonstrates that a closely related generalist wasp, Trichogramma evanescens, exploits BC in a similar way, but only after learning. Interestingly, the wasp learns to associate an H response to the odors of a mated female P. brassicae butterfly with reinforcement by parasitizing freshly laid butterfly eggs. Behavioral assays, before which we specifically inhibited long-term memory (LTM) formation with a translation inhibitor, reveal that the wasp has formed protein synthesis-dependent LTM at 24 h after learning. To our knowledge, the combination of associatively learning to exploit the sexual communication system of a host and the formation of protein synthesis-dependent LTM after a single learning event has not been documented before. We expect it to be widespread in nature, because it is highly adaptive in many species of egg parasitoids. Our finding of the exploitation of an antiaphrodisiac by multiple species of parasitic wasps suggests its use by Pieris butterflies to be under strong selective pressure.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Vinson SB. In: Insect Communication. Lewis T, editor. London: Academic; 1984. pp. 325–348.
-
- Zuk M, Kolluru GR. Exploitation of sexual signals by predators and parasitoids. Quart Rev Biol. 1998;73:415–438.
-
- Powell W. In: Pheromones of Non-Lepidopteran Insects Associated With Agricultural Plants. Hardie J, Minks AK, editors. Wallingford, United Kingdom: Centre for Agricultural Bioscience International Publishing; 1999. pp. 405–427.
-
- Fatouros NE, Dicke M, Mumm R, Meiners T, Hilker M. Foraging behavior of egg parasitoids exploiting chemical information. Behav Ecol. 2008;19:677–689.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
