

Loss of viral fitness and cross-recognition by CD8+ T cells limit HCV escape from a protective HLA-B27-restricted human immune response
- PMID: 19139562
- PMCID: PMC2631298
- DOI: 10.1172/JCI36587
Loss of viral fitness and cross-recognition by CD8+ T cells limit HCV escape from a protective HLA-B27-restricted human immune response
Abstract
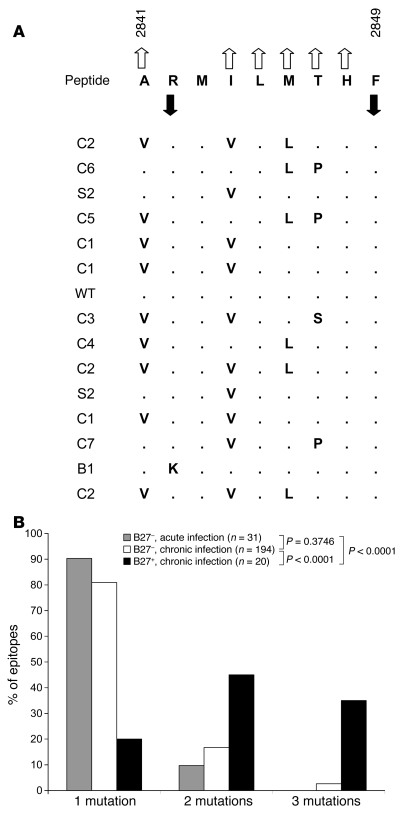
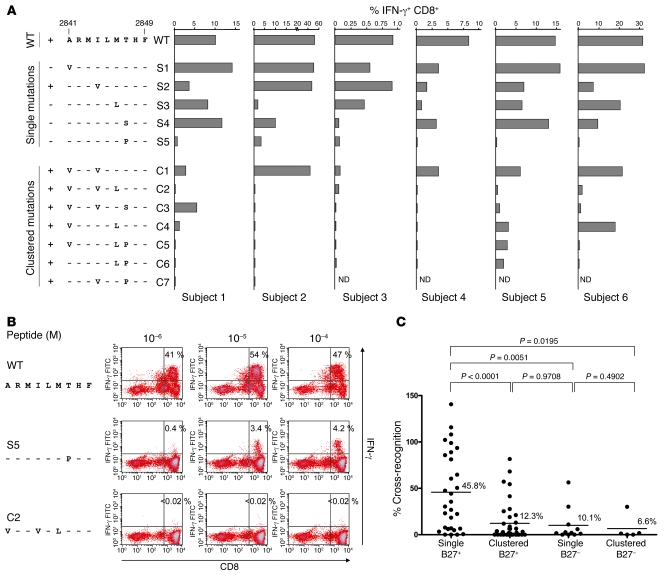
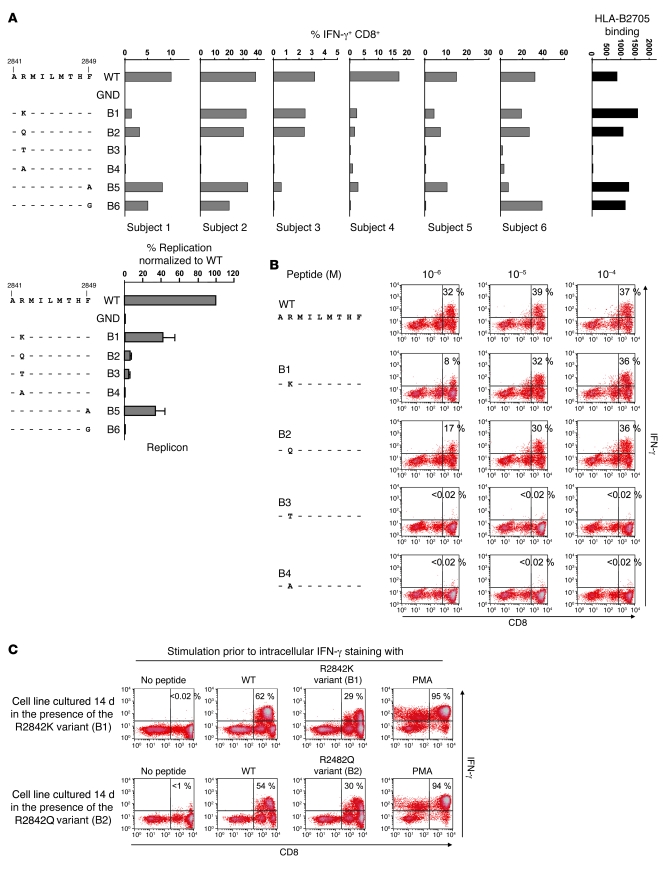
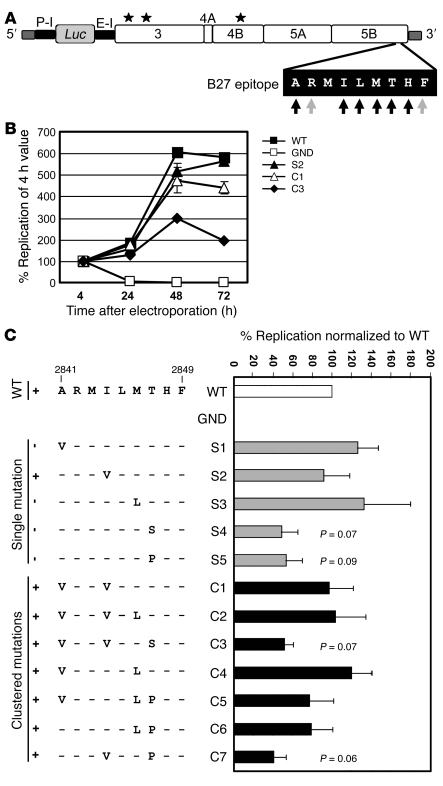
There is an association between expression of the MHC class I molecule HLA-B27 and protection following human infection with either HIV or HCV. In both cases, protection has been linked to HLA-B27 presentation of a single immunodominant viral peptide epitope to CD8+ T cells. If HIV mutates the HLA-B27-binding anchor of this epitope to escape the protective immune response, the result is a less-fit virus that requires additional compensatory clustered mutations. Here, we sought to determine whether the immunodominant HLA-B27-restricted HCV epitope was similarly constrained by analyzing the replication competence and immunogenicity of different escape mutants. Interestingly, in most HLA-B27-positive patients chronically infected with HCV, the escape mutations spared the HLA-B27-binding anchor. Instead, the escape mutations were clustered at other sites within the epitope and had only a modest impact on replication competence. Further analysis revealed that the cluster of mutations is required for efficient escape because a combination of mutations is needed to impair T cell recognition of the epitope. Artificially introduced mutations at the HLA-B27-binding anchors were found to be either completely cross-reactive or to lead to substantial loss of fitness. These results suggest that protection by HLA-B27 in HCV infection can be explained by the requirement to accumulate a cluster of mutations within the immunodominant epitope to escape T cell recognition.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
