

CPEB regulation of human cellular senescence, energy metabolism, and p53 mRNA translation
- PMID: 19141477
- PMCID: PMC2607074
- DOI: 10.1101/gad.1697808
CPEB regulation of human cellular senescence, energy metabolism, and p53 mRNA translation
Abstract
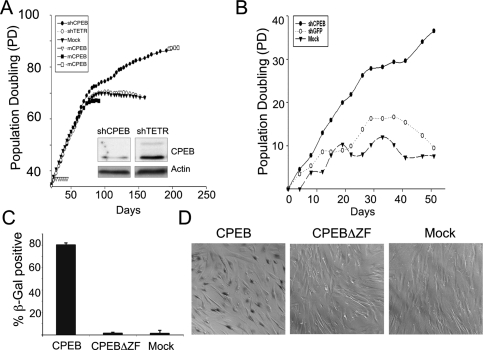
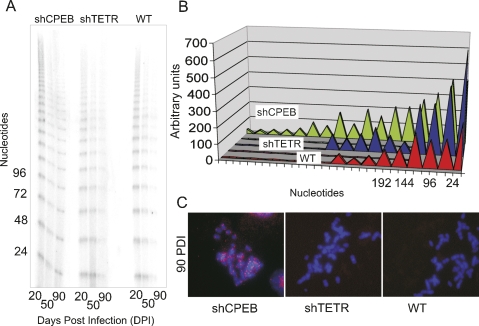
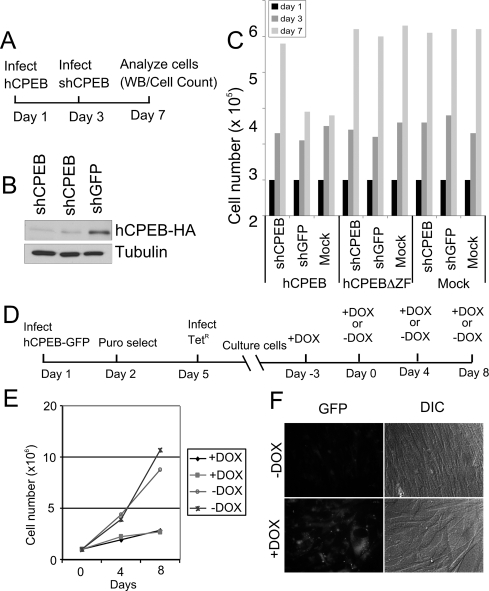
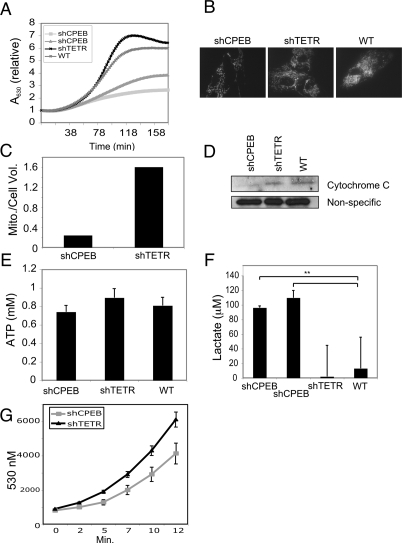
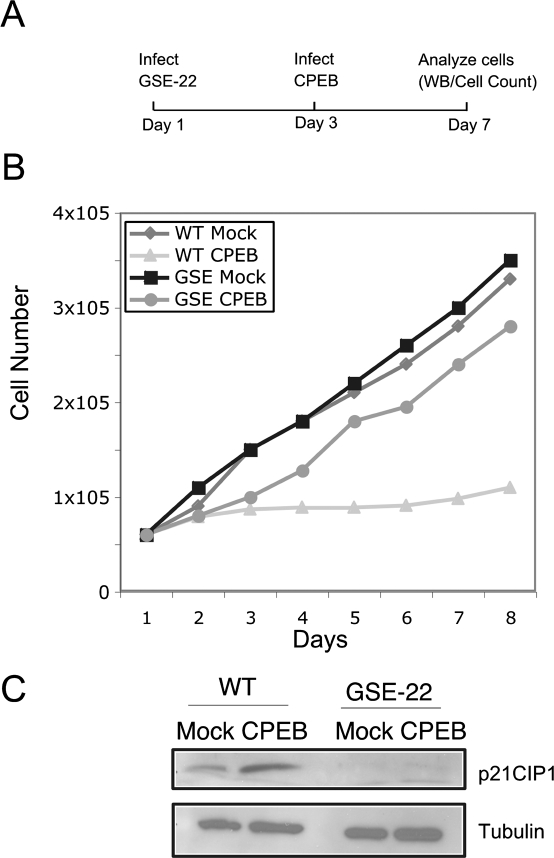
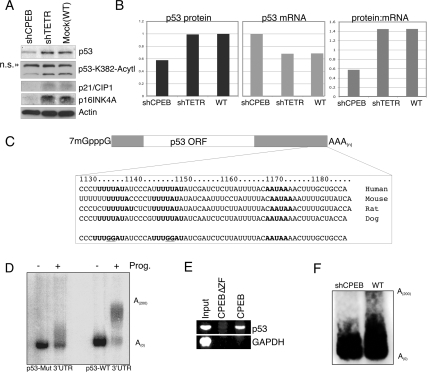
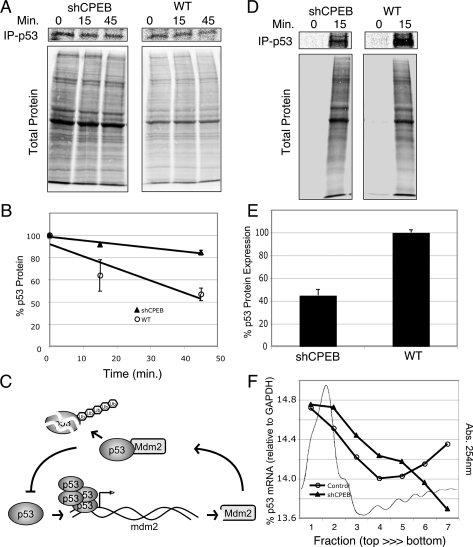
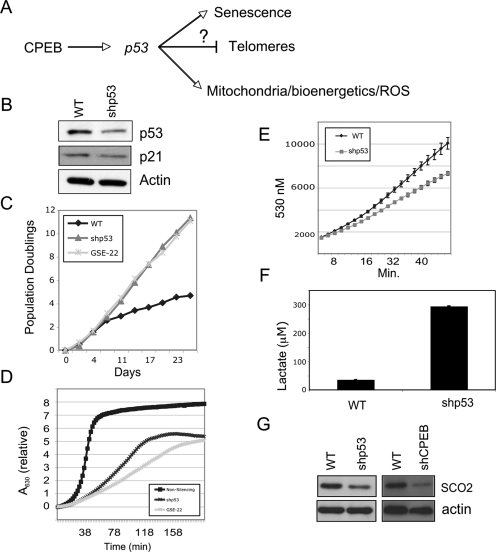
Cytoplasmic polyadenylation element-binding protein (CPEB) stimulates polyadenylation and translation in germ cells and neurons. Here, we show that CPEB-regulated translation is essential for the senescence of human diploid fibroblasts. Knockdown of CPEB causes skin and lung cells to bypass the M1 crisis stage of senescence; reintroduction of CPEB into the knockdown cells restores a senescence-like phenotype. Knockdown cells that have bypassed senescence undergo little telomere erosion. Surprisingly, knockdown of exogenous CPEB that induced a senescence-like phenotype results in the resumption of cell growth. CPEB knockdown cells have fewer mitochondria than wild-type cells and resemble transformed cells by having reduced respiration and reactive oxygen species (ROS), normal ATP levels, and enhanced rates of glycolysis. p53 mRNA contains cytoplasmic polyadenylation elements in its 3' untranslated region (UTR), which promote polyadenylation. In CPEB knockdown cells, p53 mRNA has an abnormally short poly(A) tail and a reduced translational efficiency, resulting in an approximately 50% decrease in p53 protein levels. An shRNA-directed reduction in p53 protein by about 50% also results in extended cellular life span, reduced respiration and ROS, and increased glycolysis. Together, these results suggest that CPEB controls senescence and bioenergetics in human cells at least in part by modulating p53 mRNA polyadenylation-induced translation.
Figures
References
-
- Archer H., Bar-Sagi D. Ras and Rac as activators of reactive oxygen species (ROS) Methods Mol. Biol. 2002;189:67–73. - PubMed
-
- Barnard D.C., Ryan K., Manley J.L., Richter J.D. Symplekin and xGLD-2 are required for CPEB-mediated cytoplasmic polyadenylation. Cell. 2004;119:641–651. - PubMed
-
- Ben-Porath I., Weinberg R.A. The signals and pathways activating cellular senescence. Int. J. Biochem. Cell Biol. 2005;37:961–976. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous