

doi: 10.1242/dev.015974.
Is left-right asymmetry a form of planar cell polarity?
Affiliations
- PMID: 19141667
- PMCID: PMC2687587
- DOI: 10.1242/dev.015974
Item in Clipboard
Is left-right asymmetry a form of planar cell polarity?
Development.
2009 Feb.
Abstract
Consistent left-right (LR) patterning is a clinically important embryonic process. However, key questions remain about the origin of asymmetry and its amplification across cell fields. Planar cell polarity (PCP) solves a similar morphogenetic problem, and although core PCP proteins have yet to be implicated in embryonic LR asymmetry, studies of mutations affecting planar polarity, together with exciting new data in cell and developmental biology, provide a new perspective on LR patterning. Here we propose testable models for the hypothesis that LR asymmetry propagates as a type of PCP that imposes coherent orientation onto cell fields, and that the cue that orients this polarization is a chiral intracellular structure.
Figures
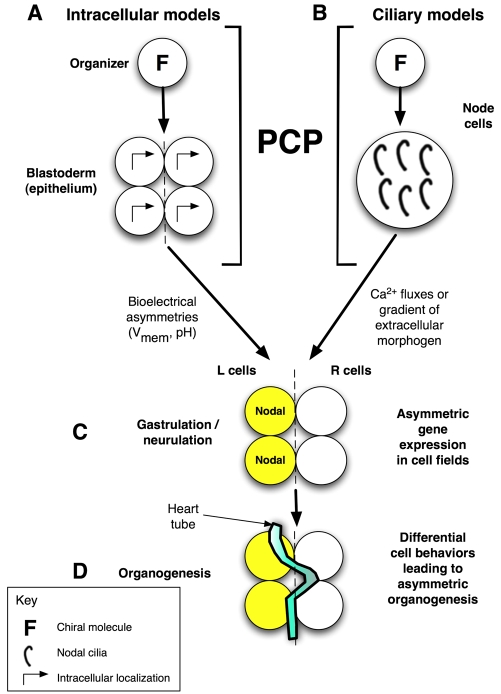
Overview of LR patterning phases and proposed role for PCP.
Consistent LR asymmetry includes sequential phases of symmetry breaking
(providing cells with a consistent orientation along the LR axis), asymmetric
gene expression, and organogenesis. (A) In our model, the initial
orientation occurs via a cytoskeletal structure (F) in very early blastomeres
that we term the LR `organizer'. This involves the coordination of the
apical-basal, LR and dorsoventral (DV) axes and is imposed across the
embryonic epithelial field by a PCP mechanism
(Aw et al., 2008). (B)
An alternative cilia-based model is also compatible with this hypothesis as
PCP is required to orient the cilia, the chiral flow of which has been
proposed to initiate asymmetry (Brueckner,
2001). In the intracellular model (A), the directional information
is converted to LR position relative to the midline by the establishment of
physiological asymmetries in membrane voltage (Vmem) that
redistribute intracellular LR morphogens
(Fukumoto et al., 2005). In
the ciliary models (B), LR position is dictated to cells by the redistribution
of an extracellular signal (e.g. Ca2+). In both models, these steps
are followed by (C) cascades of asymmetric gene expression that
(D) drive organogenesis. L, left; R, right.
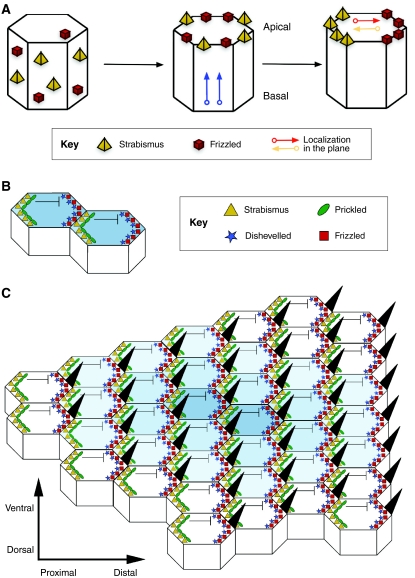
PCP establishment in the Drosophila wing. (A)
Polarity establishment in the Drosophila wing epithelium as a classic
model of PCP. PCP establishment first requires key proteins [e.g. Strabismus
(Stbm) and Frizzled (Fz)] to be apically distributed and then localized to the
proximal and distal sides of the apical surface, respectively, in the plane
that lies orthogonal to the apical-basal axis. See Zallen
(Zallen, 2007). (B)
Stbm then recruits Prickled to the proximal side of the cell, which
antagonizes Dishevelled (Dsh), hence restricting the location of the Dsh-Fz
complex to the opposite, distal side. At the same time, interactions between
the extracellular domains of Stbm and Fz at adjacent proximal and distal
membranes of neighboring cells stabilize the polarized configurations,
allowing a small clone of cells to be polarized. (C) A speculative
model of PCP (see Tree et al.,
2002b), in which the organized asymmetric distribution of core PCP
proteins spreads from a small group of polarized organizer cells. A wave of
polarized interactions emanates from an initial site of polarization outwards
in all directions through the plane of the tissue, establishing a coordinated
array of polarized cells (indicated here by the decreasing intensity of blue
shading from the center to outer cells). See Jones and Chen
(Jones and Chen, 2007).
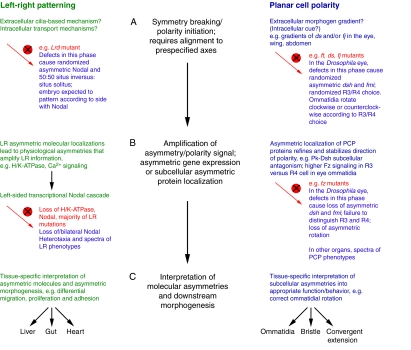
LR patterning and PCP steps share similar logic. Three phases of
patterning required for LR asymmetry (left, green) and PCP (right, blue). The
mutant vertebrate LR and Drosophila PCP eye phenotypes that result
from disruption (red circle with a cross) of each phase are described. For
simplicity, we focus on phenotypes that result from mutations in PCP genes in
the ommatidia of the Drosophila eye. (The details of analogous
phenotypes between the phases of LR and PCP establishment might differ in
other planar-polarized epithelia.) (A) In the first phase, symmetry
breaking and polarity initiation occur. Defects in this phase abolish the
directional cue and lead to random selection of polarity direction, resulting
in mutants that retain asymmetric, but randomly oriented, expression of key
downstream genes such as Nodal and fz. Mutants develop with
randomized asymmetry/polarity that follows from the direction of asymmetric
gene expression. For example, 50:50 situs inversus:situs solitus in LR
left-right dynein (Lrd) mutant or randomized
clockwise/counterclockwise rotation of ommatidia in PCP fat
(ft), dachsous (ds) and four-jointed
(fj) mutants. (B) In the second phase, the asymmetry/polarity
cue is amplified and refined over the cell field. In LR patterning, this might
occur via asymmetric ion flux or movement of extracellular morphogens by cilia
that are ultimately transduced into the left-sided Nodal
transcriptional cascade. In PCP, this occurs via the asymmetric subcellular
localization of PCP proteins. Mutations in this phase often cause loss of
asymmetric gene expression and a spectrum of LR and PCP phenotypes [loss of,
or bilateral, Nodal expression and heterotaxia in LR patterning, or
loss of asymmetric Frizzled (Fz) and loss of asymmetric rotation in ommatidia,
respectively]. (C) In the final phase, the molecular asymmetries are
differentially interpreted in the individual tissues to produce the required
morphologies. Mechanisms and phenotypes are reviewed elsewhere
(Levin, 2006;
Seifert and Mlodzik, 2007;
Tree et al., 2002a;
Wang and Nathans, 2007;
Zallen, 2007). pk,
prickled; dsh, dishevelled; fmi, flamingo; R,
photoreceptor.
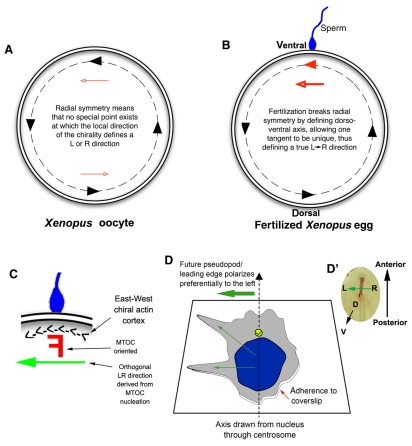
Innate chirality allows polarity determination in single cells.
(A) A Xenopus egg viewed from the animal pole; the
animal-vegetal axis lies perpendicular to the plane of the page. A consistent
`East-West' or counterclockwise chirality has been identified in the actin
cytoskeleton around the egg's periphery (black dashed line)
(Danilchik et al., 2006),
which provides different LR directional cues at distinct tangent points (black
arrowheads along the dashed line). These cues offer no unique LR orientation
because each point is equivalent to the others (red arrowheads show that cues
point rightwards on one side and leftwards on the opposite side). (B)
At fertilization, sperm entry breaks the radial symmetry and determines a
specific point on the circumference through which the midline axis of
bilateral symmetry passes. The chiral orientation of the actin cytoskeleton at
this point converts the bilateral symmetry into LR asymmetry through a linear
cue (red arrowhead defined by sperm entry) along the LR axis. Thus,
circumferential chirality can be converted into an organism-wide linear
directionality (LR) once the DV axis is determined. (C) Sperm entry
point magnified, where a putative chiral `F-molecule'
(Brown and Wolpert, 1990) in
the microtubule-organizing center (MTOC) can be oriented with respect to the
actin cortex and nucleate microtubule transport paths that have a true LR
directionality. (D) Intrinsic polarity of a differentiated HL60 cell in
culture (Xu et al., 2007).
Cells extend pseudopodia preferentially to the left of an arrow pointing from
the nucleus to the centrosome (yellow circle), revealing how HL60 cells are
intrinsically chiral, utilizing cytoplasmic structures and a polarized axis
(adhesion to coverslip versus free medium) to consistently orient the LR axis.
(D′) A chick embryo blastoderm is analogous to the cell in D, in
that its DV axis is fixed and a leftward signal must be determined to
establish sonic hedgehog (Shh) expression on the left side of the node
(brown).
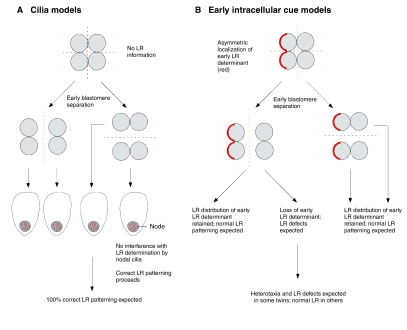
Prediction by intracellular early models of LR patterning of asymmetry
phenotypes in embryo-splitting experiments. (A) If asymmetry is
initiated by the action of cilia during gastrulation, very early blastomere
separation should result in no loss of LR information and in normal nodal
cilia. The resulting embryos are expected to exhibit 100% correct LR
patterning. (B) In intracellular models, LR information provided by
early, asymmetrically localized determinants is lost or altered in some
daughter blastomeres upon early splitting, leading to the prediction of LR
patterning defects, as is observed in human monozygotic twins and in
experiments in amphibians.
References
-
- Adler, P. N. (2002). Planar signaling and morphogenesis in Drosophila. Dev. Cell 2, 525-535. - PubMed
-
- Adler, P. N., Charlton, J. and Liu, J. (1998). Mutations in the cadherin superfamily member gene dachsous cause a tissue polarity phenotype by altering frizzled signaling. Development 125, 959-968. - PubMed
-
- Adler, P. N., Taylor, J. and Charlton, J. (2000). The domineering non-autonomy of frizzled and van Gogh clones in the Drosophila wing is a consequence of a disruption in local signaling. Mech. Dev. 96, 197-207. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
