

Review
doi: 10.1038/nrn2463.
The genetics of early telencephalon patterning: some assembly required
Affiliations
- PMID: 19143049
- PMCID: PMC2669317
- DOI: 10.1038/nrn2463
Item in Clipboard
Review
The genetics of early telencephalon patterning: some assembly required
Nat Rev Neurosci.
2008 Sep.
Abstract
The immense range of human behaviours is rooted in the complex neural networks of the cerebrum. The creation of these networks depends on the precise integration of specific neuronal subtypes that are born in different regions of the telencephalon. Here, using the mouse as a model system, we review how these proliferative zones are established. Moreover, we discuss how these regions can be traced back in development to the function of a few key genes, including those that encode fibroblast growth factors (FGFs), sonic hedgehog (SHH), bone morphogenetic proteins (BMPs), forkhead box G1 (FOXG1), paired box 6 (PAX6) and LIM homeobox protein 2 (LHX2), that pattern the early telencephalon.
Figures
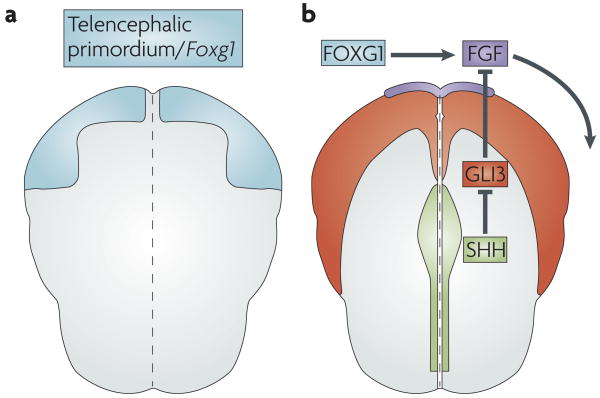
Schematics of the anterior neural plate in a mouse embryo (dorsal view, five somites stage, anterior is up). a | The region that will become the telencephalon is defined by expression of forkhead box G1 (Foxg1) (shown in blue). b | FoxG1 and sonic hedgehog (SHH; green) promote fibroblast growth factor (FGF; purple) expression in the anterior neural ridge. This patterns the nascent telencephalon (indicated by the curved arrow). SHH promotes FGF expression indirectly by inhibiting the repressor activity of GLI3 (expression of GL13 is shown in red). Consequently, SHH promotes the formation of a ventral telencephalic subdivision by inhibiting the dorsalizing effects of GLI3.
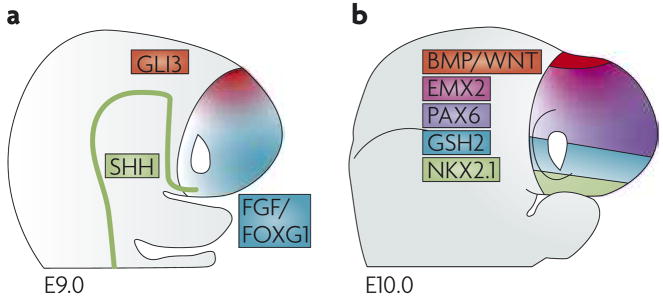
The dorsal and ventral subdivisions of the embryonic mouse telencephalon at embryonic day (E) 9.0 (a), and the four broad subdivisions at E10.0 (b). In both schematics, dorsal is up, ventral is down. The Gli3-expressing dorsal region at E9.0 is split, by E10.0, into a bone morphogenetic protein (BMP)- and Wingless/Int protein (WNT)-expressing medial region and a more lateral cortical region that expresses countergradients of empty spiracles homeobox 2 (Emx2) and paired box 6 (Pax6). The ventral region at E9.0 is split, by E10.0, into medial Nkx2.1-expressing domains and lateral Gsh2-expressing domains. At E10.0 the expression domain of Gsh2 overlaps with that of Nkx2.1; this is not represented for the sake of illustrative simplicity. Similarly, sonic hedgehog (Shh), fibroblast growth factor (FGF) and forkhead box G1 (Foxg1) expression at E10.0 is omitted.
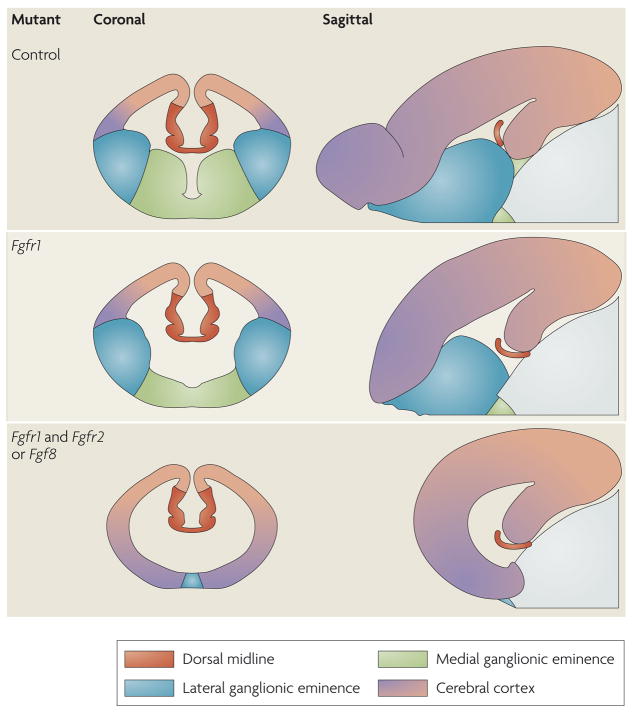
Schematics of coronal and sagittal sections in the telencephalon between embryonic day (E) 12.5 and E15.5 in various mutant mice. The colours represent areas that contain cells with different telencephalic identities. Progressively deleting more fibroblast growth factor receptor (FGFR) genes specifically in the anterior neural plate leads to diminished FGF signalling and gradually more severe truncations of telencephalic regions. Anterior ventral regions are lost first, followed by posterior dorsal regions,,.
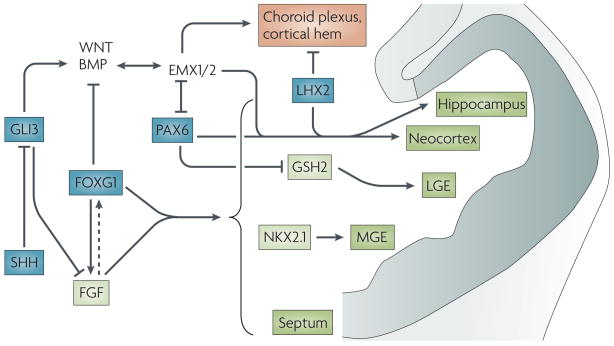
Factors that act early to establish broad telencephalic regions are shown in blue. Sonic hedgehog (SHH) ventralizes the telencephalon by antagonizing the dorsalizing effect of GLI3. By repressing Gli3, SHH, together with forkhead box G1 (FoxG1), activates fibroblast growth factor (FGF) expression. FGF might feedback and promote Foxg1 expression (dotted arrow). FoxG1 and FGF signalling are necessary for forming all regions of the telencephalon (shown in green), except for the dorsomedial region (shown in orange). Downstream transcription factors, such as GSH2 and NKX2.1, then form specific subdivisions. In the dorsal telencephalon, GLI3’s promotion of the expression of bone morphogenetic proteins (BMPs) and Wingless/Int proteins (WNTs) is required for EMX-gene expression. The products of the EMx genes, along with PAX6 and LHX2, further subdivide the dorsal telencephalon. LGE, lateral ganglionic eminence; MGE, medial ganglionic eminence.
References
-
- Beddington RS, Robertson EJ. Anterior patterning in mouse. Trends Genet. 1998;14:277–284. - PubMed
-
- Wilson SW, Rubenstein JL. Induction and dorsoventral patterning of the telencephalon. Neuron. 2000;28:641–651. - PubMed
-
- Rallu M, Corbin JG, Fishell G. Parsing the prosencephalon. Nature Rev Neurosci. 2002;3:5031–5040. - PubMed
-
- Ingham P, McMahon AP. Hedgehog signaling in animal development: paradigms and principles. Genes Dev. 2001;15:3059–3087. - PubMed
-
- Kobayashi D, et al. Early subdivisions in the neural plate define distinct competence for inductive signals. Development. 2002;129:83–93. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
