

Experimental evidence for ascorbate-dependent electron transport in leaves with inactive oxygen-evolving complexes
- PMID: 19144767
- PMCID: PMC2649403
- DOI: 10.1104/pp.108.132621
Experimental evidence for ascorbate-dependent electron transport in leaves with inactive oxygen-evolving complexes
Abstract
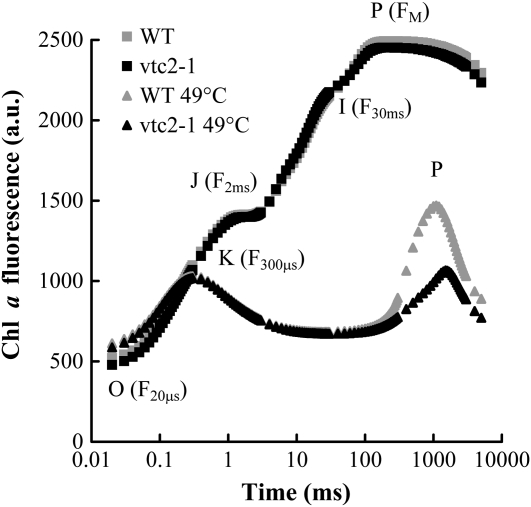
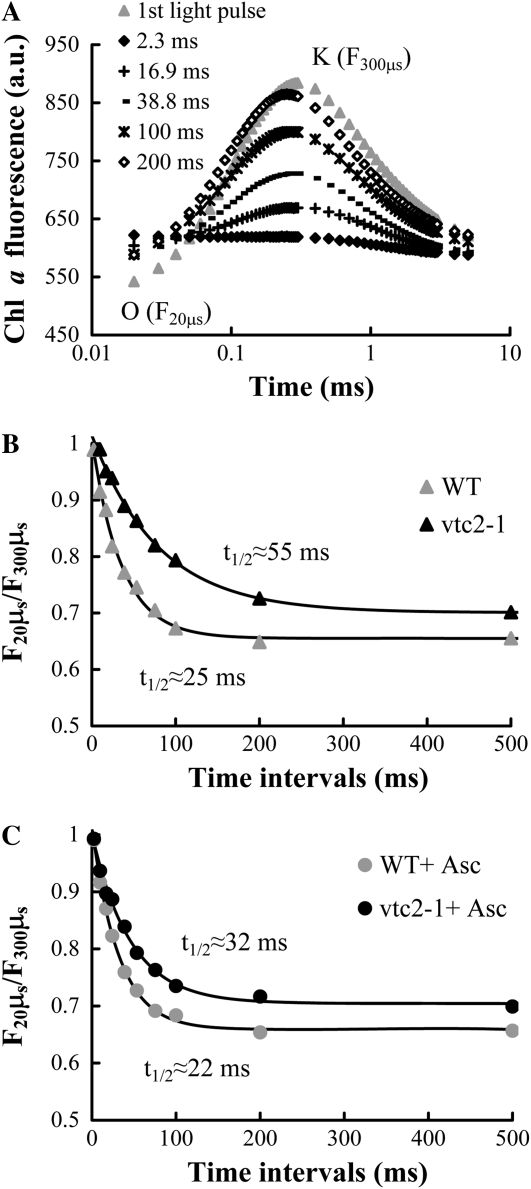
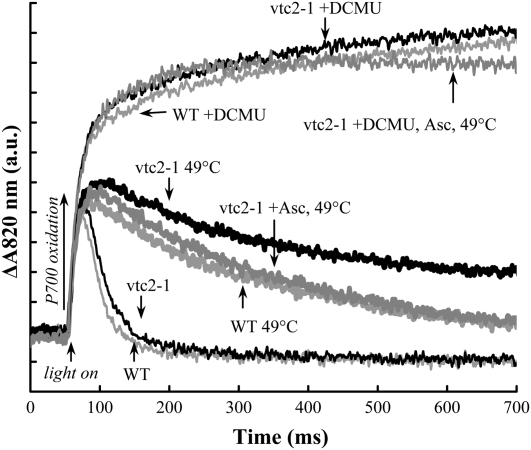
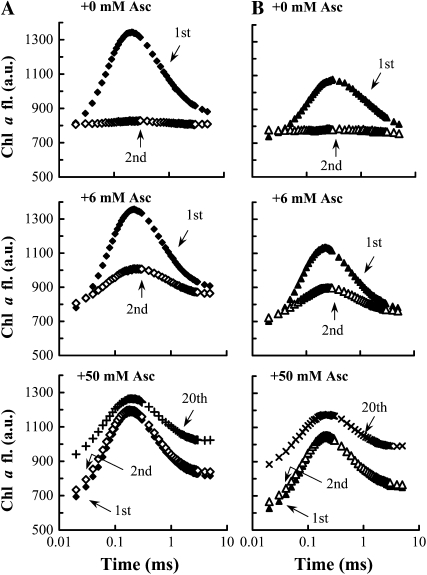
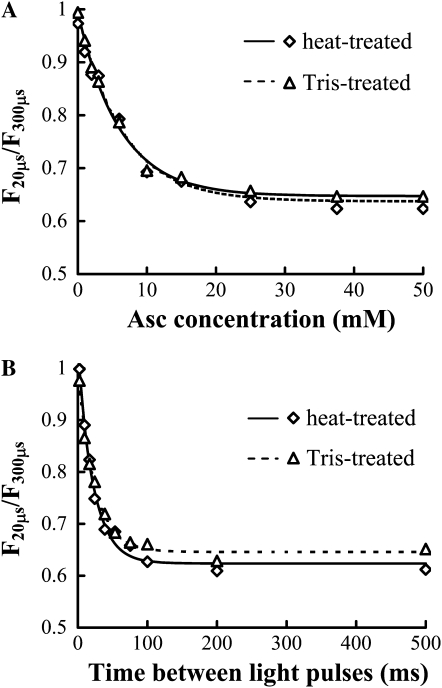
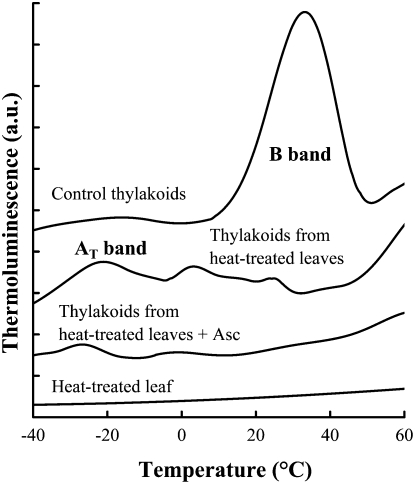
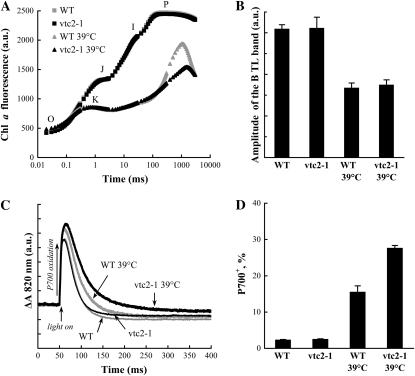
Previously, we showed that in barley (Hordeum vulgare) leaves with heat-inactivated oxygen-evolving complexes, photosystem II (PSII) has access to a large pool of alternative electron donors. Based on in vitro data, we proposed that this donor was ascorbate, yet this hypothesis has not been substantiated in vivo. In this paper, with the aid of chlorophyll a fluorescence induced by short (5-ms) light pulses and 820-nm absorbance transient measurements on wild-type and ascorbate-deficient (vtc2-1) mutant leaves of Arabidopsis (Arabidopsis thaliana), we show that in heat-treated leaves the rate of electron donation to PSII as well as the 3-(3,4-dichlorophenyl)-1,1-dimethylurea-sensitive electron transport toward photosystem I depend on the ascorbate content of the leaves: upon ascorbate treatment, the donation half-time in the wild type and the mutant decreased from 25 to 22 ms and from 55 to 32 ms, respectively. Thermoluminescence measurements show that Tyr(Z)(+) is involved in the electron transfer from ascorbate to PSII. These data and the similar ascorbate dependencies of the heat-treated and the tris(hydroxymethyl)aminomethane-treated thylakoid membranes, with maximal donation half-times of about 16 ms, show that ascorbate is capable of supporting a sustained electron transport activity in leaves containing inactivated oxygen-evolving complexes. This alternative electron transport appears to be ubiquitous in the plant kingdom and is present in the green alga Chlamydomonas reinhardtii, and its rate depends on the physiological state of the plants and on environmental conditions. Our data suggest that ascorbate, as an alternative PSII electron donor, plays a physiological role in heat-stressed plants.
Figures
References
-
- Babcock GT, Blankenship RE, Sauer K (1976) Reaction kinetics for positive charge accumulation on the water side of chloroplast photosystem II. FEBS Lett 61 286–289 - PubMed
-
- Barra M, Haumann M, Dau H (2005) Specific loss of the extrinsic 18 kDa protein from photosystem II upon heating to 47°C causes inactivation of oxygen evolution likely due to Ca release from the Mn-complex. Photosynth Res 84 231–237 - PubMed
-
- Blubaugh DJ, Atamian M, Babcock GT, Golbeck JH, Cheniae GM (1991) Photoinhibition of hydroxylamine-extracted photosystem II membranes: identification of the sites of photodamage. Biochemistry 30 7586–7597 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
