

Interactions between prefrontal cortex and cerebellum revealed by trace eyelid conditioning
- PMID: 19144967
- PMCID: PMC2632850
- DOI: 10.1101/lm.1178309
Interactions between prefrontal cortex and cerebellum revealed by trace eyelid conditioning
Abstract
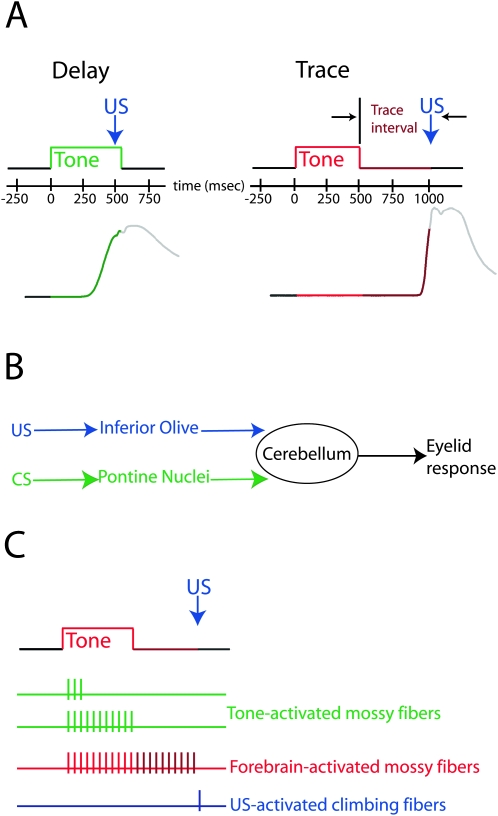
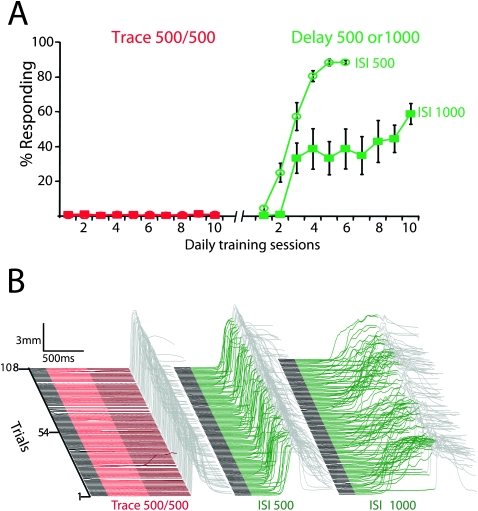
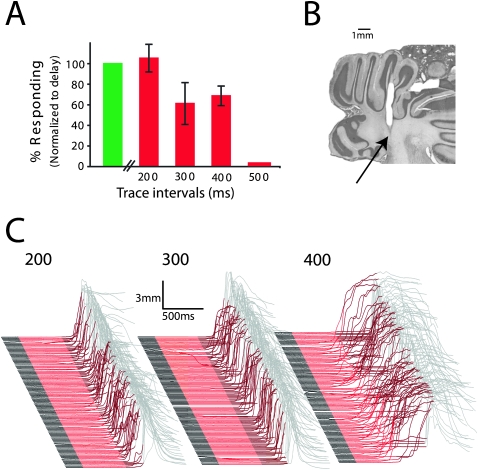
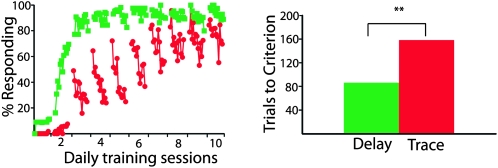
Eyelid conditioning has proven useful for analysis of learning and computation in the cerebellum. Two variants, delay and trace conditioning, differ only by the relative timing of the training stimuli. Despite the subtlety of this difference, trace eyelid conditioning is prevented by lesions of the cerebellum, hippocampus, or medial prefrontal cortex (mPFC), whereas delay eyelid conditioning is prevented by cerebellar lesions and is largely unaffected by forebrain lesions. Here we test whether these lesion results can be explained by two assertions: (1) Cerebellar learning requires temporal overlap between the mossy fiber inputs activated by the tone conditioned stimulus (CS) and the climbing fiber inputs activated by the reinforcing unconditioned stimulus (US), and therefore (2) trace conditioning requires activity that outlasts the presentation of the CS in a subset of mossy fibers separate from those activated directly by the CS. By use of electrical stimulation of mossy fibers as a CS, we show that cerebellar learning during trace eyelid conditioning requires an input that persists during the stimulus-free trace interval. By use of reversible inactivation experiments, we provide evidence that this input arises from the mPFC and arrives at the cerebellum via a previously unidentified site in the pontine nuclei. In light of previous PFC recordings in various species, we suggest that trace eyelid conditioning involves an interaction between the persistent activity of delay cells in mPFC-a putative mechanism of working memory-and motor learning in the cerebellum.
Figures
References
-
- Aitkin L.M., Boyd J. Acoustic input to the lateral pontine nuclei. Hear. Res. 1978;1:67–77. - PubMed
-
- Bao S., Chen L., Thompson R.F. Learning- and cerebellum-dependent neuronal activity in the lateral pontine nucleus. Behav. Neurosci. 2000;114:254–261. - PubMed
-
- Beylin A.V., Gandhi C.C., Wood G.E., Talk A.C., Matzel L.D., Shors T.J. The role of the hippocampus in trace conditioning: Temporal discontinuity or task difficulty? Neurobiol. Learn. Mem. 2001;76:447–461. - PubMed
-
- Bodner M., Kroger J., Fuster J.M. Auditory memory cells in dorsolateral prefrontal cortex. Neuroreport. 1996;7:1905–1908. - PubMed
-
- Boyd J., Aitkin L.M. Responses of single units in the pontine nuclei of the cat to acoustic stimulation. Neurosci. Lett. 1976;3:259–263. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous