

Two tumor suppressors, p27Kip1 and patched-1, collaborate to prevent medulloblastoma
- PMID: 19147535
- PMCID: PMC2637533
- DOI: 10.1158/1541-7786.MCR-08-0369
Two tumor suppressors, p27Kip1 and patched-1, collaborate to prevent medulloblastoma
Abstract
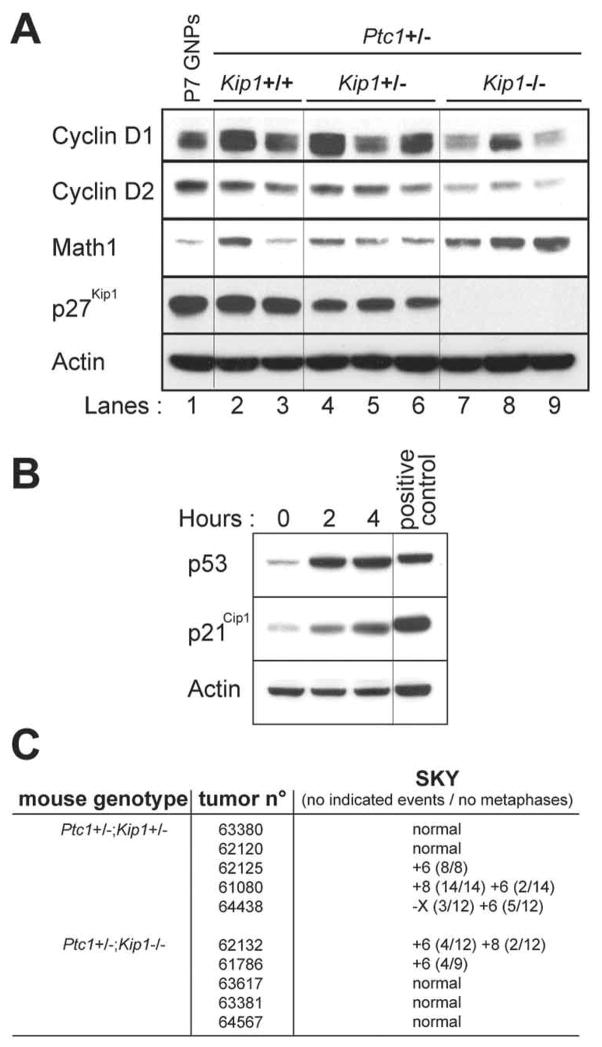
Two cyclin-dependent kinase inhibitors, p18(Ink4c) and p27(Kip1), are required for proper cerebellar development. Loss of either of these proteins conferred a proliferative advantage to granule neuron progenitors, although inactivation of Kip1 exerted a greater effect. Mice heterozygous for Patched-1 (Ptc1+/-) that are either heterozygous or nullizygous for Kip1 developed medulloblastoma rapidly and with high penetrance. All tumors from Ptc1+/-;Kip1+/- or Ptc1+/-;Kip1-/- mice failed to express the wild-type Ptc1 allele, consistent with its role as a canonical "two-hit" tumor suppressor. In contrast, expression of the wild-type p27(Kip1) protein was invariably maintained in medulloblastomas arising in Ptc1+/-;Kip1+/- mice, indicating that Kip1 is haploinsufficient for tumor suppression. Although medulloblastomas occurring in Ptc1+/- mice were histopathologically heterogeneous and contained intermixed regions of both rapidly proliferating and nondividing more differentiated cells, tumors that also lacked Kip1 were uniformly less differentiated, more highly proliferative, and invasive. Molecular analysis showed that the latter medulloblastomas exhibited constitutive activation of the Sonic hedgehog signaling pathway without loss of functional p53. Apart from gains or losses of single chromosomes, with gain of chromosome 6 being the most frequent, no other chromosomal anomalies were identified by spectral karyotyping, and half of the medulloblastomas so examined retained a normal karyotype. In this respect, this mouse medulloblastoma model recapitulates the vast majority of human medulloblastomas that do not sustain TP53 mutations and are not aneuploid.
Figures



References
-
- Marino S. Medulloblastoma: developmental mechanisms out of control. Trends Mol Med. 2005;11:17–22. - PubMed
-
- Wang VY, Zoghbi HY. Genetic regulation of cerebellar development. Nat Rev Neurosci. 2001;27:484–91. - PubMed
-
- Sherr C, Roberts JM. CDK inhibitors: positive and negative regulators of G1-phase progression. Genes Dev. 1999;13:1501–12. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
