

Review
doi: 10.1038/nrmicro2070.
Pyroptosis: host cell death and inflammation
Affiliations
- PMID: 19148178
- PMCID: PMC2910423
- DOI: 10.1038/nrmicro2070
Item in Clipboard
Review
Pyroptosis: host cell death and inflammation
Nat Rev Microbiol.
2009 Feb.
Abstract
Eukaryotic cells can initiate several distinct programmes of self-destruction, and the nature of the cell death process (non-inflammatory or proinflammatory) instructs responses of neighbouring cells, which in turn dictates important systemic physiological outcomes. Pyroptosis, or caspase 1-dependent cell death, is inherently inflammatory, is triggered by various pathological stimuli, such as stroke, heart attack or cancer, and is crucial for controlling microbial infections. Pathogens have evolved mechanisms to inhibit pyroptosis, enhancing their ability to persist and cause disease. Ultimately, there is a competition between host and pathogen to regulate pyroptosis, and the outcome dictates life or death of the host.
Figures
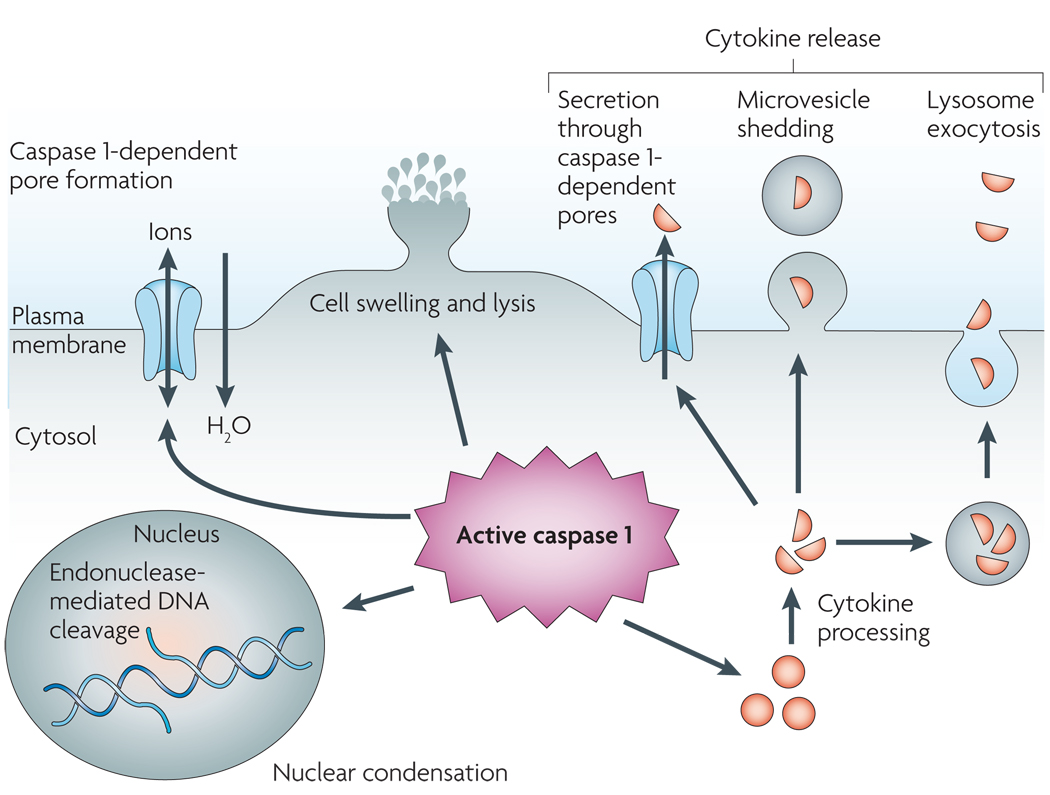
Caspase 1 is cleaved and activated in response to multiple stimuli, but once activated, caspase 1 results in a conserved programme of cell death referred to as pyroptosis. Caspase 1 activation also leads to rapid formation of plasma-membrane pores with a diameter of 1.1–2.4 nm. These pores dissipate cellular ionic gradients, allowing water influx, cell swelling and osmotic lysis. The pro-forms of interleukin-1β (IL-1β) and IL-18 are processed by caspase 1 and released during pyroptosis, although the exact mechanism of secretion remains controversial. Secretion does not require lysis and is temporally associated with caspase 1-dependent pore formation, suggesting that these pores facilitate cytokine release. Other suggested secretion mechanisms include caspase 1-independent lysosome exocytosis and microvesicle shedding. Caspase 1 activity results in cleavage of chromosomal DNA by an unidentified endonuclease. Cleavage of DNA does not result in the oligonucleosomal fragments observed during apoptosis. Nuclear condensation is also observed but nuclear integrity is maintained, unlike the nuclear fragmentation observed during apoptosis.
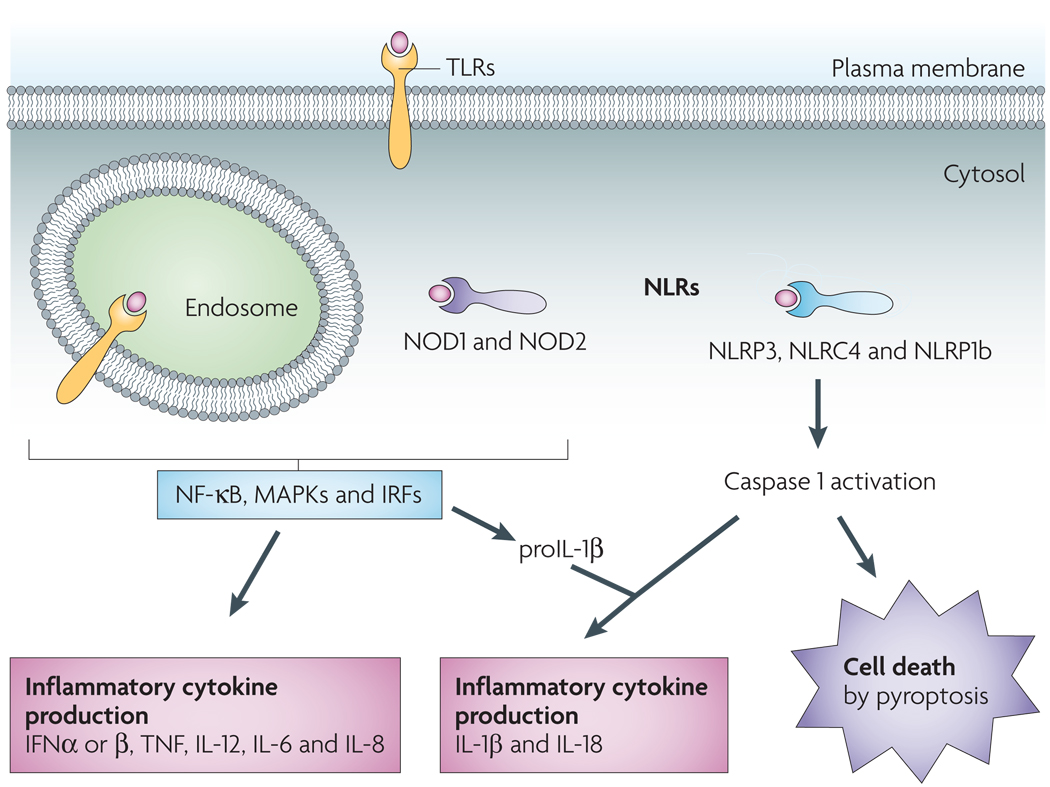
Leucine-rich repeat (LRR) domains mediate host recognition of pathogen- and danger-associated molecular patterns. Toll-like receptors (TLRs) are LRR-containing transmembrane proteins that detect danger signals located in the extracellular milieu and within endosomes. TLRs initiate a signalling cascade that leads to cellular activation (through nuclear factor-κB (NF-κB)-, mitogen-activated protein kinase (MAPK)- and interferon (IFN)-regulatory factor (IRF)-dependent pathways) and inflammatory cytokine production (including IFNα, IFNβ, tumour necrosis factor (TNF), interleukin-12 (IL-12), IL-6, IL-8 and pro-IL-1β). Nod-like receptors (NLRs) function in the recognition of danger signals introduced into the host cell cytosol. Like TLRs, NOD1 (nucleotide-binding oligomerization domain-containing protein 1) and NOD2 stimulation results in cytokine production. Another subset of NLRs mediate activation of the cysteine protease caspase 1, which triggers caspase 1-dependent cell death (pyroptosis) and processing and release of the inflammatory cytokines IL-18 and IL-1β. NLRC4, NLR family CARD domain-containing protein 4; NLRP3, NACHT, LRR and PYD domains-containing protein 3. NLRP1b, NAHCT, LRR and PYD domains-containing protein 1b.
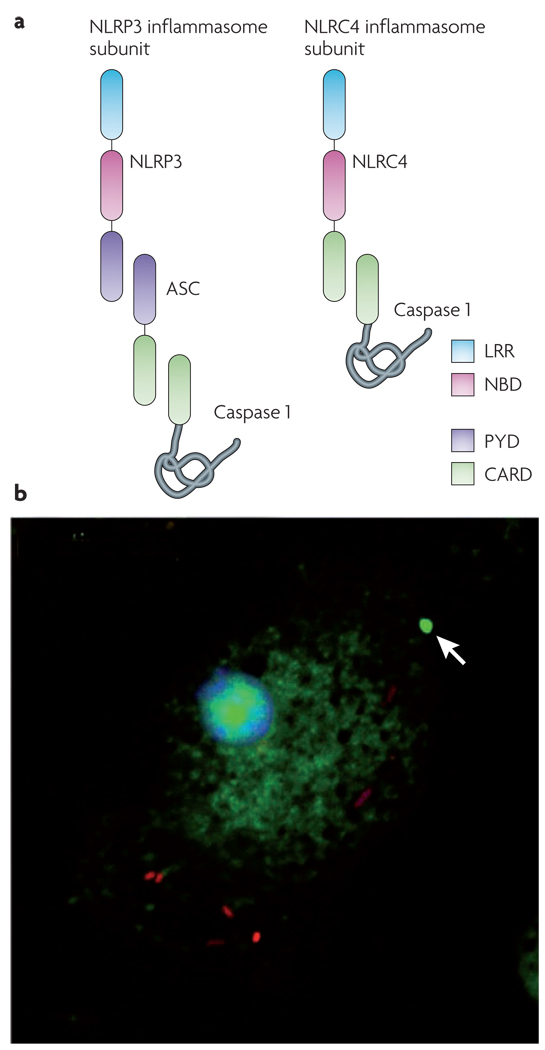
a | Nod-like receptor (NLR) leucine-rich repeat (LRR) domains are implicated in sensing a range of intracellular ‘danger’ signals. After ligand recognition, the nucleotide-binding and oligomerization domain (NBD) mediates nucleotide-dependent self-association of NLRs. Some NLRs, such as NLRP3 (NACHT, LRR and PYD domains-containing protein 3; also called NALP3), contain a pyrin (PYD) domain that interacts with the adapter protein ASC (apoptosis-associated speck-like protein containing a CARD). ASC contains a caspase activation and recruitment domain (CARD) that binds and facilitates activation of caspase 1. Other NLRs, such as NLRC4 (NLR family CARD domain-containing protein 4; also known as IPAF), contain a CARD and can directly interact with caspase 1. However, ASC is often required for NLRC4-dependent caspase 1 activation, indicating that ASC may participate in NLRC4 inflammasome formation or play an additional part in caspase 1 activation. b | Salmonella (red) infection of macrophages results in activation of caspase 1 (green), which is visualized here using a fluorescently labelled inhibitor of the active enzyme. Active caspase 1 is often concentrated within a single focus (indicated by the arrow) and diffusely distributed throughout the cytoplasm. A similar distribution of active caspase 1 is seen in macrophages treated with Bacillus anthracis lethal toxin.

Caspase 1 plays a protective part in the response to microbial infection. a | In response to infection, quiescent cells undergo caspase 1 activation and pyroptosis, allowing cleavage and release of interleukin-18 (IL-18), IL-1β and other inflammatory intracellular contents. Quiescent cells can also undergo ‘activation’ in response to inflammatory mediators, thereby lowering the threshold for caspase 1 activation and pyroptosis and stimulating increased production of IL-1β. b | As infection progresses, the inflammation that occurs as a consequence of pyroptosis leads to an increased population of activated cells that are primed to undergo pyroptosis and have increased inflammatory potential. c | Inflammatory contents produced during pyroptosis recruit and activate immune cells and stimulate the development of adaptive immune responses. This contributes to the control and ultimate resolution of microbial infection, and returns tissues to their resting state. Alternatively, caspase 1 activation can be detrimental, as mutations in Nod-like receptor (NLR) proteins or the persistence of sterile inflammatory stimuli can result in inappropriate and/or excessive caspase 1 activation. The inflammation produced by this process increases the population of activated cells that are primed to undergo pyroptosis and express increased levels of IL-1β, and the amplification cycle persists (b). This potentiates the response and maintains an inflammatory state, which, if uninterrupted, leads to pathology.
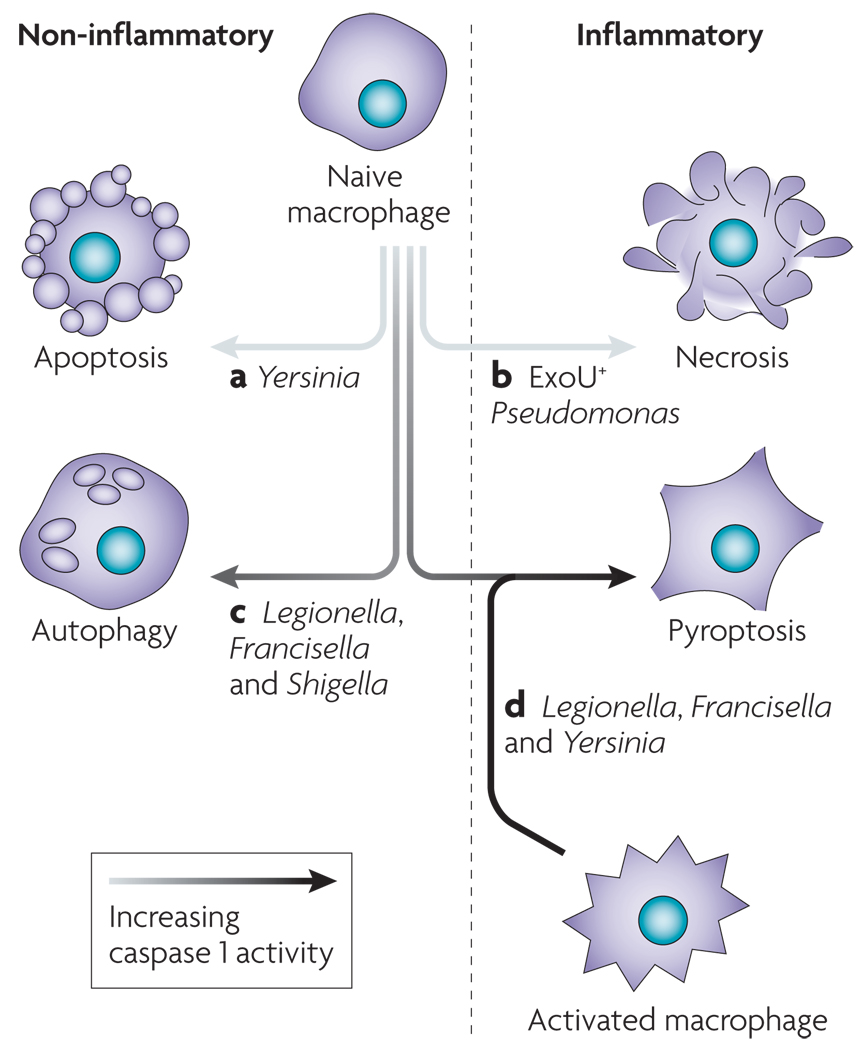
Pathogens have mechanisms for modulating cell death by inhibiting caspase 1 activation or inducing an alternative form of cell death that is more conducive to their continued replication. Yersinia (a) and Pseudomonas (b) translocate type III secretion effectors, resulting in apoptosis and necrosis, respectively. Pathogens can fail to induce robust caspase 1 activation owing to suboptimal ‘danger’ signal production by the pathogen (c). In addition, host mutations may not allow sufficient levels of caspase 1 activation to trigger pyroptosis. These infected macrophages often display features that are consistent with autophagy. Robust production of caspase 1-activating ligands by Legionella during infection of a susceptible macrophage triggers pyroptosis (d). Not all cells are uniformly susceptible to pyroptosis, and macrophage activation enhances caspase 1 activation (FIG. 4) in response to Yersinia and Francisella infection, which do not stimulate pyroptosis in naive macrophages.
References
-
- Samali A, Zhivotovsky B, Jones D, Nagata S, Orrenius S. Apoptosis: cell death defined by caspase activation. Cell Death Differ. 1999;6:495–496. - PubMed
-
- Albert ML. Death-defying immunity: do apoptotic cells influence antigen processing and presentation? Nature Rev. Immunol. 2004;4:223–231. - PubMed
-
- Frantz S, et al. Targeted deletion of caspase-1 reduces early mortality and left ventricular dilatation following myocardial infarction. J. Mol. Cell. Cardiol. 2003;35:685–694. - PubMed
-
- Fantuzzi G, Dinarello CA. Interleukin-18 and interleukin-1 beta: two cytokine substrates for ICE (caspase-1) J. Clin. Immunol. 1999;19:1–11. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
