

PBX1 is dispensable for neural commitment of RA-treated murine ES cells
- PMID: 19148706
- PMCID: PMC2758398
- DOI: 10.1007/s11626-008-9162-5
PBX1 is dispensable for neural commitment of RA-treated murine ES cells
Abstract
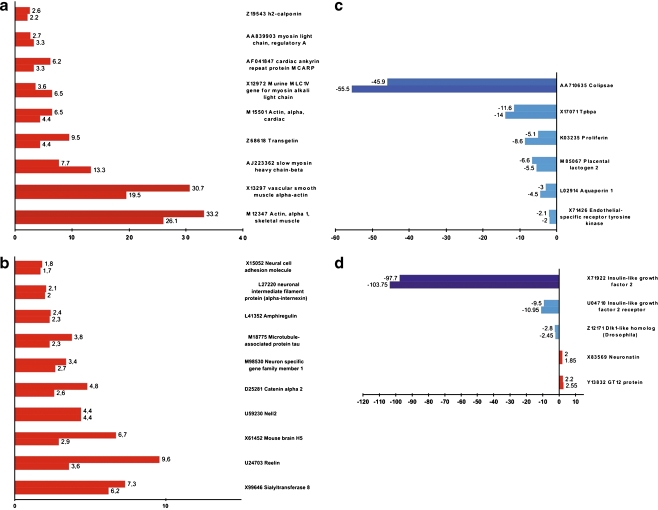
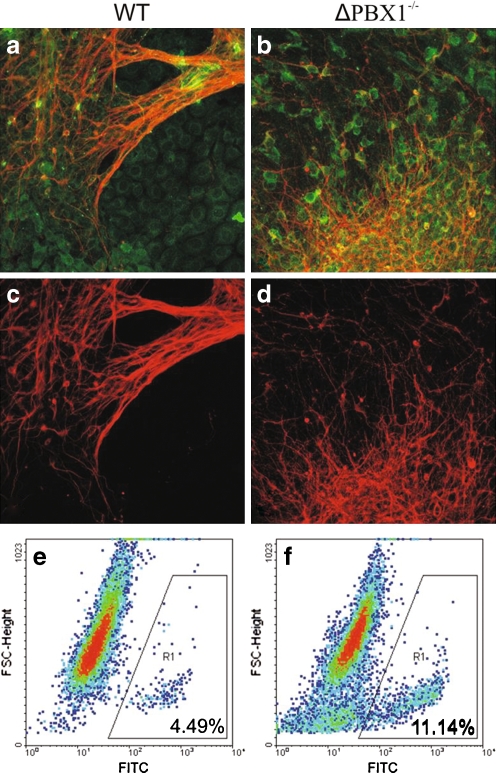
Experimentation with PBX1 knockout mice has shown that PBX1 is necessary for early embryogenesis. Despite broad insight into PBX1 function, little is known about the underlying target gene regulation. Utilizing the Cre-loxP system, we targeted a functionally important part of the homeodomain of PBX1 through homozygous deletion of exon-6 and flanking intronic regions leading to exon 7 skipping in embryonic stem (ES) cells. We induced in vitro differentiation of wild-type and PBX1 mutant ES cells by aggregation and retinoic acid (RA) treatment and compared their profiles of gene expression at the ninth day post-reattachment to adhesive media. Our results indicate that PBX1 interactions with HOX proteins and DNA are dispensable for RA-induced ability of ES to express neural genes and point to a possible involvement of PBX1 in the regulation of imprinted genes.
Figures





References
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'PubMed', 'value': '12376109', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/12376109/'}]}
- Adamson S. L.; Lu Y.; Whiteley K. J.; Holmyard D.; Hemberger M.; Pfarrer C.; Cross J. C. Interactions between trophoblast cells and the maternal and fetal circulation in the mouse placenta. Dev. Biol. 250: 358–373; 2002. - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1016/j.ydbio.2003.09.041', 'is_inner': False, 'url': 'https://doi.org/10.1016/j.ydbio.2003.09.041'}, {'type': 'PubMed', 'value': '14732407', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/14732407/'}]}
- Ahn J. I.; Lee K. H.; Shin D. M.; Shim J. W.; Lee J. S.; Chang S. Y.; Lee Y. S.; Brownstein M. J.; Lee S. H. Comprehensive transcriptome analysis of differentiation of embryonic stem cells into midbrain and hindbrain neurons. Dev. Biol. 265: 491–501; 2004. doi:10.1016/j.ydbio.2003.09.041. - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1016/S0169-328X(03)00256-0', 'is_inner': False, 'url': 'https://doi.org/10.1016/s0169-328x(03)00256-0'}, {'type': 'PubMed', 'value': '12941464', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/12941464/'}]}
- Aihara K.; Kuroda S.; Kanayama N.; Matsuyama S.; Tanizawa K.; Horie M. A neuron-specific EGF family protein, NELL2, promotes survival of neurons through mitogen-activated protein kinases. Brain Res. Mol. Brain Res. 116: 86–93; 2003. doi:10.1016/S0169-328X(03)00256-0. - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1006/dbio.1995.1085', 'is_inner': False, 'url': 'https://doi.org/10.1006/dbio.1995.1085'}, {'type': 'PubMed', 'value': '7729574', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/7729574/'}]}
- Bain G.; Kitchens D.; Yao M.; Huettner J. E.; Gottlieb D. I. Embryonic stem cells express neuronal properties in vitro. Dev. Biol. 168: 342–357; 1995. doi:10.1006/dbio.1995.1085. - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'PubMed', 'value': '2598811', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/2598811/'}]}
- Beddington R. S.; Robertson E. J. An assessment of the developmental potential of embryonic stem cells in the midgestation mouse embryo. Development 105: 733–737; 1989. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
