

New genes of Xanthomonas citri subsp. citri involved in pathogenesis and adaptation revealed by a transposon-based mutant library
- PMID: 19149882
- PMCID: PMC2653504
- DOI: 10.1186/1471-2180-9-12
New genes of Xanthomonas citri subsp. citri involved in pathogenesis and adaptation revealed by a transposon-based mutant library
Abstract
Background: Citrus canker is a disease caused by the phytopathogens Xanthomonas citri subsp. citri, Xanthomonas fuscans subsp. aurantifolli and Xanthomonas alfalfae subsp. citrumelonis. The first of the three species, which causes citrus bacterial canker type A, is the most widely spread and severe, attacking all citrus species. In Brazil, this species is the most important, being found in practically all areas where citrus canker has been detected. Like most phytobacterioses, there is no efficient way to control citrus canker. Considering the importance of the disease worldwide, investigation is needed to accurately detect which genes are related to the pathogen-host adaptation process and which are associated with pathogenesis.
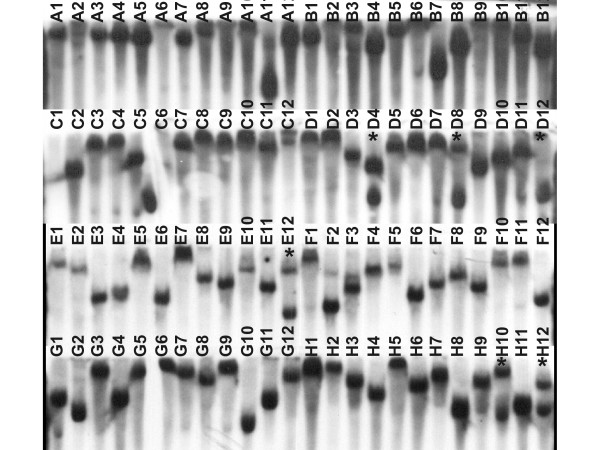
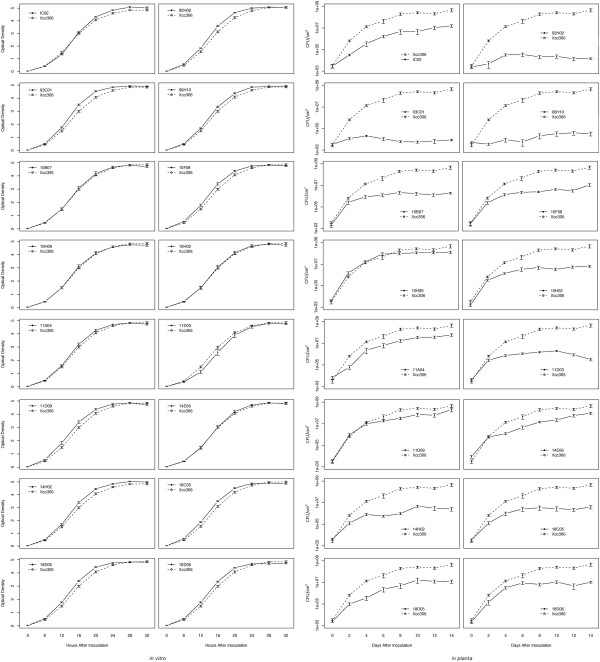
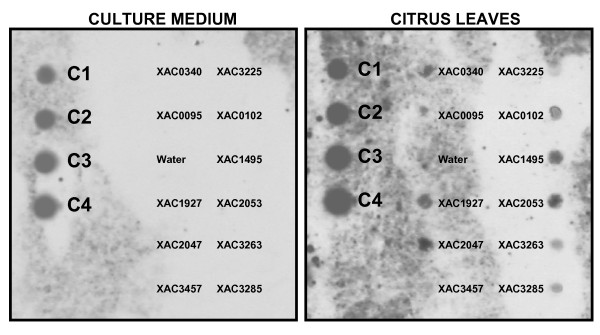
Results: Through transposon insertion mutagenesis, 10,000 mutants of Xanthomonas citri subsp. citri strain 306 (Xcc) were obtained, and 3,300 were inoculated in Rangpur lime (Citrus limonia) leaves. Their ability to cause citrus canker was analyzed every 3 days until 21 days after inoculation; a set of 44 mutants showed altered virulence, with 8 presenting a complete loss of causing citrus canker symptoms. Sequencing of the insertion site in all 44 mutants revealed that 35 different ORFs were hit, since some ORFs were hit in more than one mutant, with mutants for the same ORF presenting the same phenotype. An analysis of these ORFs showed that some encoded genes were previously known as related to pathogenicity in phytobacteria and, more interestingly, revealed new genes never implicated with Xanthomonas pathogenicity before, including hypothetical ORFs. Among the 8 mutants with no canker symptoms are the hrpB4 and hrpX genes, two genes that belong to type III secretion system (TTSS), two hypothetical ORFS and, surprisingly, the htrA gene, a gene reported as involved with the virulence process in animal-pathogenic bacteria but not described as involved in phytobacteria virulence. Nucleic acid hybridization using labeled cDNA probes showed that some of the mutated genes are differentially expressed when the bacterium is grown in citrus leaves. Finally, comparative genomic analysis revealed that 5 mutated ORFs are in new putative pathogenicity islands.
Conclusion: The identification of these new genes related with Xcc infection and virulence is a great step towards the understanding of plant-pathogen interactions and could allow the development of strategies to control citrus canker.
Figures
References
-
- Whiteside J, Garnsey S, Timmer L. Compendium of citrus diseases. Saint Paul: APS Press; 1988.
-
- Feichtenberger E. In: Anais do V Seminário Internacional de Citros – Tratos Culturais. Donadio L, editor. Bebedouro: Fundação Cargill; 1998. Manejo ecológico das principais doenças fúngicas e bacterianas dos citros no Brasil; p. 517.
-
- da Silva ACR, Ferro JA, Reinach FC, Farah CS, Furlan LR, Quaggio RB, Monteiro-Vitorello CB, Sluys MAV, Almeida NF, Alves LMC, do Amaral AM, Bertolini MC, Camargo LEA, Camarotte G, Cannavan F, Cardozo J, Chambergo F, Ciapina LP, Cicarelli RMB, Coutinho LL, Cursino-Santos JR, El-Dorry H, Faria JB, Ferreira AJS, Ferreira RCC, Ferro MIT, Formighieri EF, Franco MC, Greggio CC, Gruber A, Katsuyama AM, Kishi LT, Leite RP, Lemos EGM, Lemos MVF, Locali EC, Machado MA, Madeira AMBN, Martinez-Rossi NM, Martins EC, Meidanis J, Menck CFM, Miyaki CY, Moon DH, Moreira LM, Novo MTM, Okura VK, Oliveira MC, Oliveira VR, Pereira HA, Rossi A, Sena JAD, Silva C, de Souza RF, Spinola LAF, Takita MA, Tamura RE, Teixeira EC, Tezza RID, dos Santos MT, Truffi D, Tsai SM, White FF, Setubal JC, Kitajima JP. Comparison of the genomes of two Xanthomonas pathogens with differing host specificities. Nature. 2002;417(6887):459–463. doi: 10.1038/417459a. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
