

Information theoretical quantification of cooperativity in signalling complexes
- PMID: 19149897
- PMCID: PMC2637831
- DOI: 10.1186/1752-0509-3-9
Information theoretical quantification of cooperativity in signalling complexes
Abstract
Background: Intra-cellular information exchange, propelled by cascades of interacting signalling proteins, is essential for the proper functioning and survival of cells. Now that the interactome of several organisms is being mapped and several structural mechanisms of cooperativity at the molecular level in proteins have been elucidated, the formalization of this fundamental quantity, i.e. information, in these very diverse biological contexts becomes feasible.
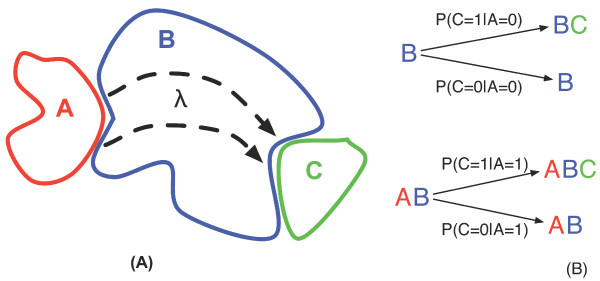
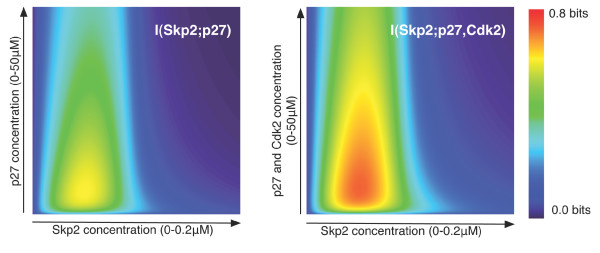
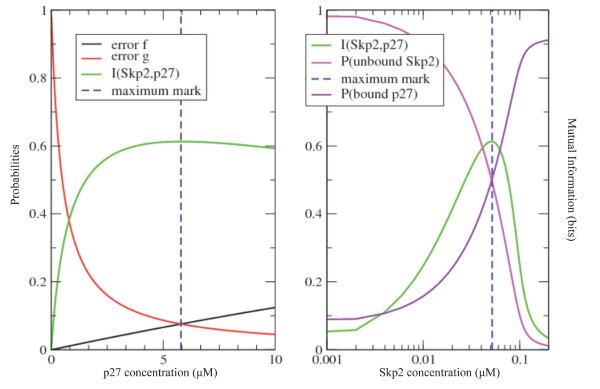
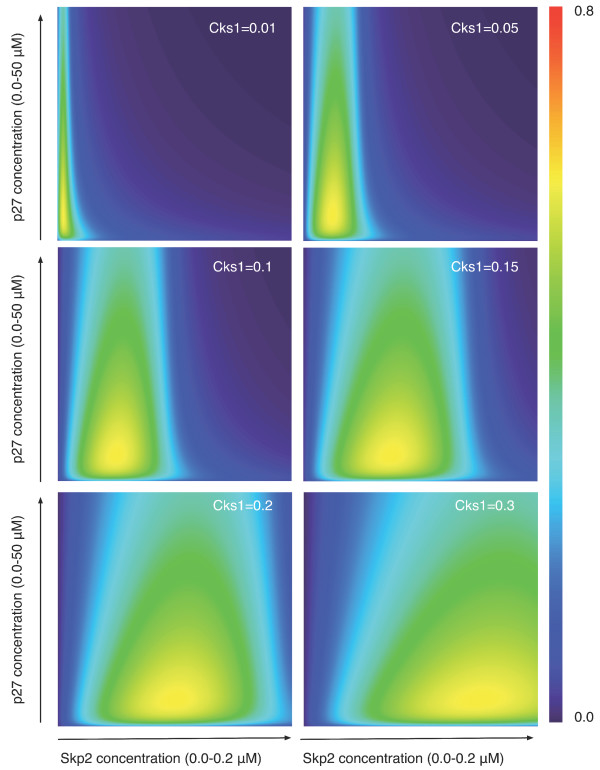
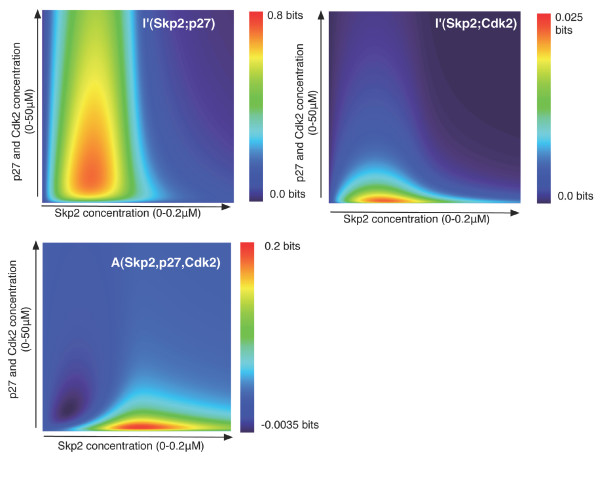
Results: We show here that Shannon's mutual information quantifies information in biological system and more specifically the cooperativity inherent to the assembly of macromolecular complexes. We show how protein complexes can be considered as particular instances of noisy communication channels. Further we show, using a portion of the p27 regulatory pathway, how classical equilibrium thermodynamic quantities such as binding affinities and chemical potentials can be used to quantify information exchange but also to determine engineering properties such as channel noise and channel capacity. As such, this information measure identifies and quantifies those protein concentrations that render the biochemical system most effective in switching between the active and inactive state of the intracellular process.
Conclusion: The proposed framework provides a new and original approach to analyse the effects of cooperativity in the assembly of macromolecular complexes. It shows the conditions, provided by the protein concentrations, for which a particular system acts most effectively, i.e. exchanges the most information. As such this framework opens the possibility of grasping biological qualities such as system sensitivity, robustness or plasticity directly in terms of their effect on information exchange. Although these parameters might also be derived using classical thermodynamic parameters, a recasting of biological signalling in terms of information exchange offers an alternative framework for visualising network cooperativity that might in some cases be more intuitive.
Figures
References
-
- Rousseau F, Schymkowitz J. A systems biology perspective on protein structural dynamics and signal transduction. Curr Opin Struct Biol. 2005;15:23–30. - PubMed
-
- Senear DF, Ross JB, Laue TM. Analysis of protein and DNA-mediated contributions to cooperative assembly of protein-DNA complexes. Methods. 1998;16:3–20. - PubMed
-
- Luque I, Leavitt SA, Freire E. The linkage between protein folding and functional cooperativity: two sides of the same coin? Annu Rev Biophys Biomol Struct. 2002;31:235–256. - PubMed
-
- Goh CS, Milburn D, Gerstein M. Conformational changes associated with protein-protein interactions. Curr Opin Struct Biol. 2004;14:104–109. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
