

betaTrCP- and Rsk1/2-mediated degradation of BimEL inhibits apoptosis
- PMID: 19150432
- PMCID: PMC2655121
- DOI: 10.1016/j.molcel.2008.12.020
betaTrCP- and Rsk1/2-mediated degradation of BimEL inhibits apoptosis
Abstract
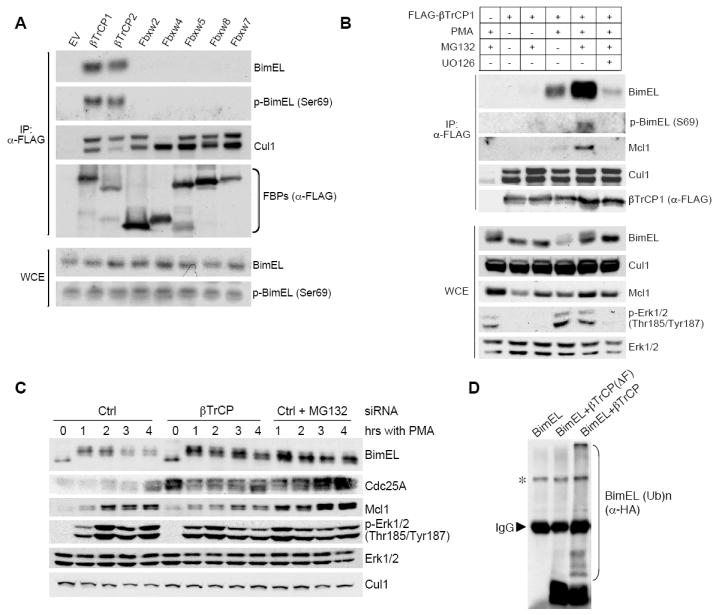
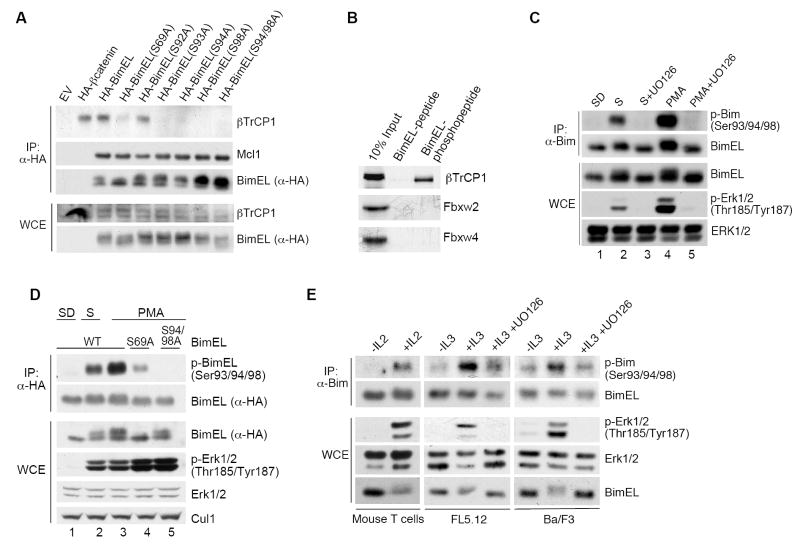
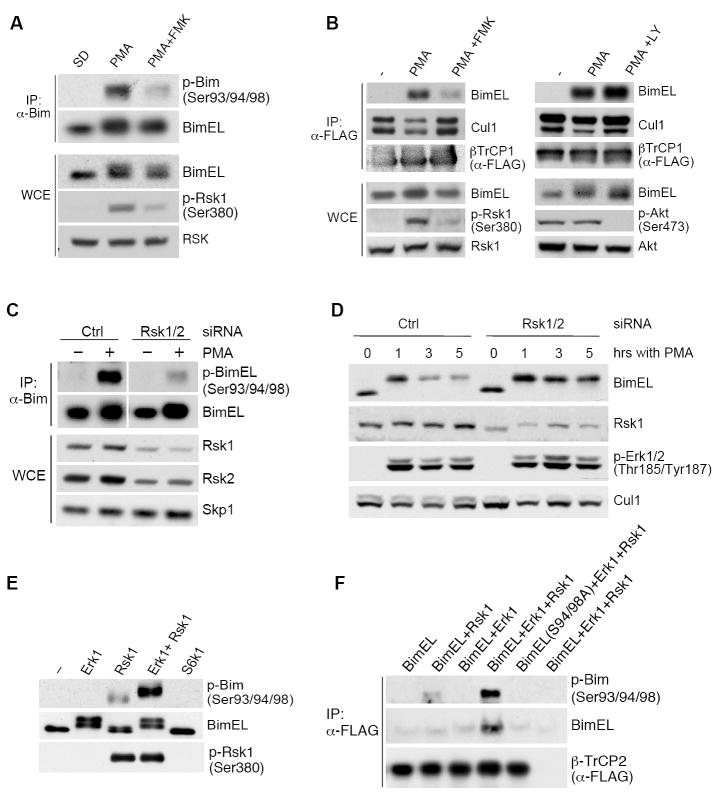
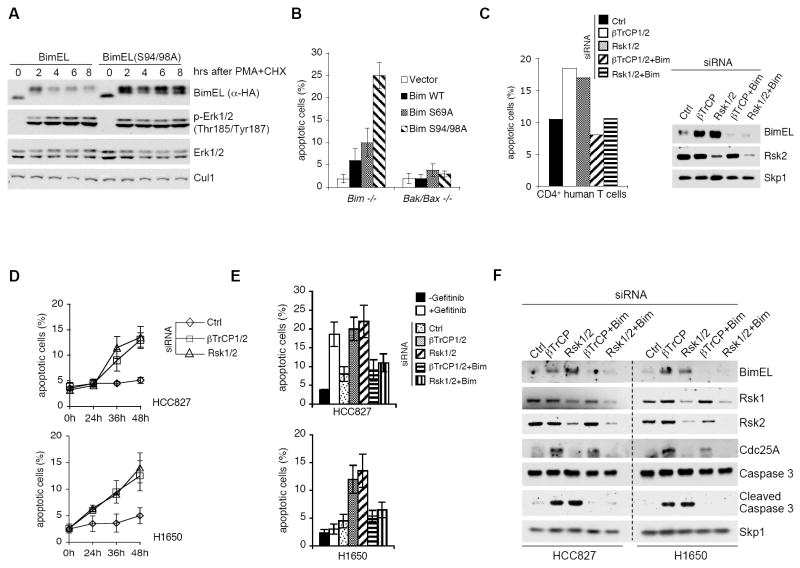
The BimEL tumor suppressor is a potent proapoptotic BH3-only protein. We found that, in response to survival signals, BimEL was rapidly phosphorylated on three serine residues in a conserved degron, facilitating binding and degradation via the F box protein betaTrCP. Phosphorylation of the BimEL degron was executed by Rsk1/2 and promoted by the Erk1/2-mediated phosphorylation of BimEL on Ser69. Compared to wild-type BimEL, a BimEL phosphorylation mutant unable to bind betaTrCP was stabilized and consequently potent at inducing apoptosis by the intrinsic mitochondrial pathway. Moreover, although non-small cell lung cancer (NSCLC) cells often become resistant to gefitinib (a clinically relevant tyrosine kinase inhibitor that induces apoptosis through BimEL), silencing of either betaTrCP or Rsk1/2 resulted in BimEL-mediated apoptosis of both gefitinib-sensitive and gefitinib-insensitive NSCLC cells. Our findings reveal that betaTrCP promotes cell survival in cooperation with the ERK-RSK pathway by targeting BimEL for degradation.
Figures
References
-
- Anjum R, Blenis J. The RSK family of kinases: emerging roles in cellular signalling. Nat Rev Mol Cell Biol. 2008;9:747–758. - PubMed
-
- Belloc F, Moreau-Gaudry F, Uhalde M, Cazalis L, Jeanneteau M, Lacombe F, Praloran V, Mahon FX. Imatinib and nilotinib induce apoptosis of chronic myeloid leukemia cells through a Bim-dependant pathway modulated by cytokines. Cancer Biol Ther. 2007;6:912–919. - PubMed
-
- Busino L, Bassermann F, Maiolica A, Lee C, Nolan PM, Godinho SI, Draetta GF, Pagano M. SCFFbxl3 controls the oscillation of the circadian clock by directing the degradation of cryptochrome proteins. Science. 2007;316:900–904. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
