

A TLR3 ligand that exhibits potent inhibition of influenza virus replication and has strong adjuvant activity has the potential for dual applications in an influenza pandemic
- PMID: 19150474
- PMCID: PMC7115584
- DOI: 10.1016/j.vaccine.2008.12.048
A TLR3 ligand that exhibits potent inhibition of influenza virus replication and has strong adjuvant activity has the potential for dual applications in an influenza pandemic
Abstract
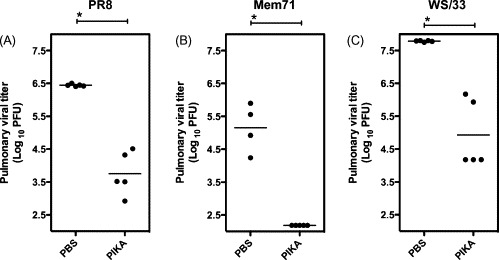
The appearance and spread of the H5N1 highly pathogenic avian influenza (HPAI) raise concern of a possible pandemic. Current preventive measures include the development of a pre-pandemic influenza vaccine and stockpiling of neuraminidase inhibitors. However, their benefits can be significantly reduced by mutations in the hemagglutinin or neuraminidase resulting in antigenic changes and the appearance of drug-resistance, respectively. Drugs that target the innate immune system to achieve a 'heightened antiviral' state represent another class of antiviral agents that could contribute to the control and treatment of influenza infection. In this study, PIKA (a stabilized dsRNA) provides broad-spectrum prophylaxis against a number of influenza A viruses. In addition, when PIKA was admixed with influenza vaccine preparations, including a formalin-inactivated whole-virion H5 vaccine, significant adjuvanting effect leading to accelerated viral clearance was observed in a murine model. These biological effects appear to be mediated by the ability of PIKA to promote the maturation of dendritic cells, including up-regulation of co-stimulatory molecules, such as CD80 and CD86, and the induction of various cytokines and chemokines. Toll-like receptor 3 (TLR3) was shown to recognize PIKA in a concentration-dependent manner. The potency and versatility in its activities make PIKA an attractive candidate for use in an influenza pandemic.
Figures






Similar articles
-
An adjuvant for the induction of potent, protective humoral responses to an H5N1 influenza virus vaccine with antigen-sparing effect in mice.J Virol. 2010 Sep;84(17):8639-49. doi: 10.1128/JVI.00596-10. Epub 2010 Jun 10. J Virol. 2010. PMID: 20538850 Free PMC article.
-
Characterization of an influenza A H5N2 reassortant as a candidate for live-attenuated and inactivated vaccines against highly pathogenic H5N1 viruses with pandemic potential.Vaccine. 2006 Nov 17;24(47-48):6859-66. doi: 10.1016/j.vaccine.2006.06.023. Epub 2006 Jun 28. Vaccine. 2006. PMID: 17050041
-
Generation of an attenuated H5N1 avian influenza virus vaccine with all eight genes from avian viruses.Vaccine. 2007 Oct 16;25(42):7379-84. doi: 10.1016/j.vaccine.2007.08.011. Epub 2007 Aug 24. Vaccine. 2007. PMID: 17870216
-
Development of a mucosal vaccine for influenza viruses: preparation for a potential influenza pandemic.Expert Rev Vaccines. 2007 Apr;6(2):193-201. doi: 10.1586/14760584.6.2.193. Expert Rev Vaccines. 2007. PMID: 17408369 Review.
-
New strategies for the development of H5N1 subtype influenza vaccines: progress and challenges.BioDrugs. 2011 Oct 1;25(5):285-98. doi: 10.1007/BF03256169. BioDrugs. 2011. PMID: 21942913 Review.
Cited by
-
TLR Agonists as Mediators of Trained Immunity: Mechanistic Insight and Immunotherapeutic Potential to Combat Infection.Front Immunol. 2021 Feb 18;11:622614. doi: 10.3389/fimmu.2020.622614. eCollection 2020. Front Immunol. 2021. PMID: 33679711 Free PMC article. Review.
-
Fast and long-lasting immune response to S-trimer COVID-19 vaccine adjuvanted by PIKA.Mol Biomed. 2021;2(1):29. doi: 10.1186/s43556-021-00054-z. Epub 2021 Sep 27. Mol Biomed. 2021. PMID: 34766005 Free PMC article.
-
Toll-like Receptor Mediation in SARS-CoV-2: A Therapeutic Approach.Int J Mol Sci. 2022 Sep 14;23(18):10716. doi: 10.3390/ijms231810716. Int J Mol Sci. 2022. PMID: 36142620 Free PMC article. Review.
-
Synergistic TLR2/6 and TLR9 activation protects mice against lethal influenza pneumonia.PLoS One. 2012;7(1):e30596. doi: 10.1371/journal.pone.0030596. Epub 2012 Jan 27. PLoS One. 2012. PMID: 22299046 Free PMC article.
-
The Innate Antiviral Response in Animals: An Evolutionary Perspective from Flagellates to Humans.Viruses. 2019 Aug 16;11(8):758. doi: 10.3390/v11080758. Viruses. 2019. PMID: 31426357 Free PMC article. Review.
References
-
- Nicholson K.G., Colegate A.E., Podda A., Stephenson I., Wood J., Ypma E. Safety and antigenicity of non-adjuvanted and MF59-adjuvanted influenza A/Duck/Singapore/97 (H5N3) vaccine: a randomised trial of two potential vaccines against H5N1 influenza. Lancet. 2001;357(June (9272)):1937–1943. - PubMed
-
- Treanor J.J., Campbell J.D., Zangwill K.M., Rowe T., Wolff M. Safety and immunogenicity of an inactivated subvirion influenza A (H5N1) vaccine. N Engl J Med. 2006;354(March (13)):1343–1351. - PubMed
-
- Bresson J.L., Perronne C., Launay O., Gerdil C., Saville M., Wood J. Safety and immunogenicity of an inactivated split-virion influenza A/Vietnam/1194/2004 (H5N1) vaccine: phase I randomised trial. Lancet. 2006;367(May (9523)):1657–1664. - PubMed
-
- Nichol K.L., Treanor J.J. Vaccines for seasonal and pandemic influenza. J Infect Dis. 2006;194(November (Suppl. 2)):S111–S118. - PubMed
-
- Stephenson I., Nicholson K.G., Colegate A., Podda A., Wood J., Ypma E. Boosting immunity to influenza H5N1 with MF59-adjuvanted H5N3 A/Duck/Singapore/97 vaccine in a primed human population. Vaccine. 2003;21(April (15)):1687–1693. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
