

Critical roles of coactivator p300 in mouse embryonic stem cell differentiation and Nanog expression
- PMID: 19150979
- PMCID: PMC2666568
- DOI: 10.1074/jbc.M805562200
Critical roles of coactivator p300 in mouse embryonic stem cell differentiation and Nanog expression
Abstract
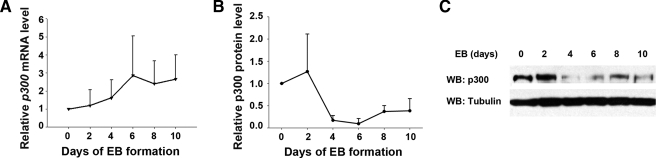
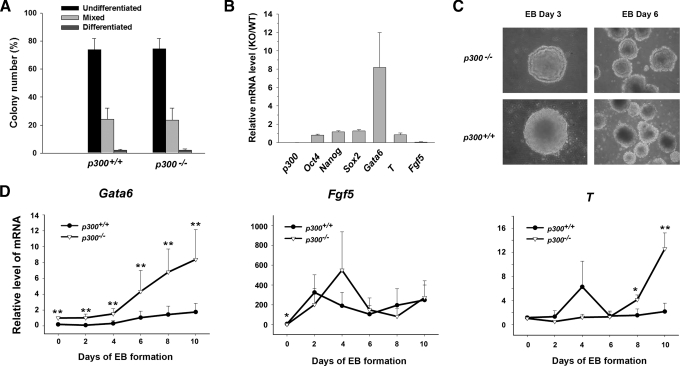
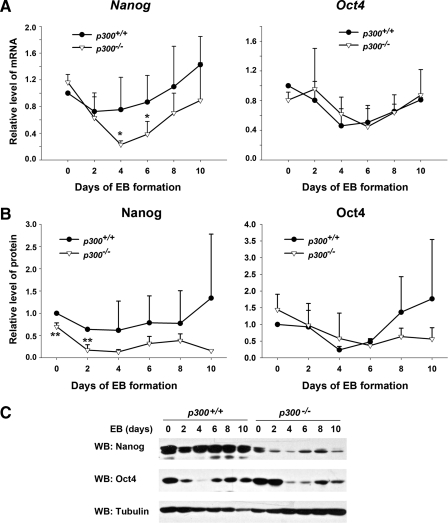
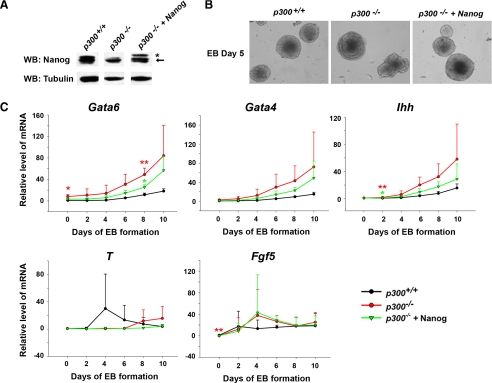
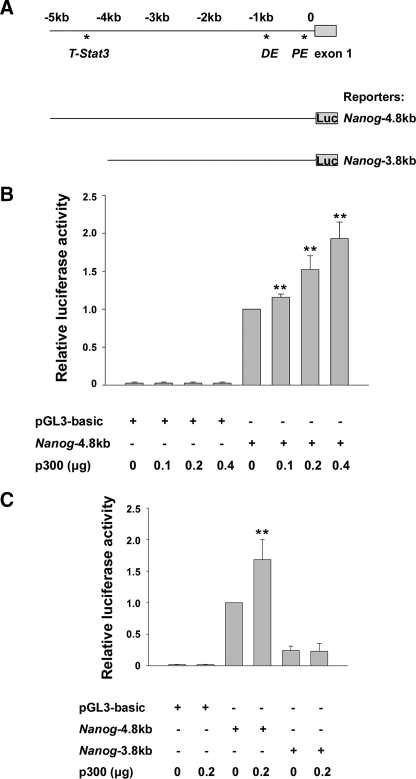
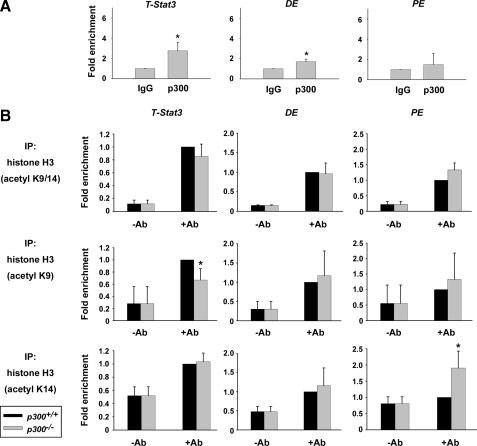
p300 is a well known histone acetyltransferase and coactivator that plays pivotal roles in many physiological processes. Despite extensive research for the functions of p300 in embryogenesis and transcription regulation, its roles in regulating embryonic stem (ES) cell pluripotency are poorly understood. To address this issue, we investigated the self-renewal ability and early differentiation process in both wild-type mouse ES cells and ES cells derived from p300 knock-out (p300(-/-)) mice. We found that p300 ablation did not affect self-renewal capacity overtly when ES cells were maintained under undifferentiated conditions. However, the absence of p300 caused a significantly abnormal expression pattern of germ layer markers when differentiation was induced by embryoid body (EB) formation. Interestingly, the expression level of pluripotency marker Nanog but not Oct4 was markedly lower in EBs from p300(-/-) ES cells compared with that in EBs from wild-type ES cells. Exogenous expression of Nanog rescued abnormal expression of extra-embryonic endoderm marker partially but not mesoderm and ectoderm markers. Furthermore, we demonstrate that p300 was directly involved in modulating Nanog expression. Importantly, epigenetic modification of histone acetylation at the distal regulatory region of Nanog was found to be dependent on the presence of p300, which could contribute to the mechanism of regulating Nanog expression by p300. Collectively, our results show that p300 plays an important role in the differentiation process of ES cells and provide the first evidence for the involvement of p300 in regulating Nanog expression during differentiation, probably through epigenetic modification of histone on Nanog.
Figures
Similar articles
-
Transcriptional reprogramming and chromatin remodeling accompanies Oct4 and Nanog silencing in mouse trophoblast lineage.Stem Cells Dev. 2014 Feb 1;23(3):219-29. doi: 10.1089/scd.2013.0328. Epub 2013 Nov 7. Stem Cells Dev. 2014. PMID: 24059348 Free PMC article.
-
The histone acetyltransferase Myst2 regulates Nanog expression, and is involved in maintaining pluripotency and self-renewal of embryonic stem cells.FEBS Lett. 2015 Apr 2;589(8):941-50. doi: 10.1016/j.febslet.2015.02.029. Epub 2015 Mar 3. FEBS Lett. 2015. PMID: 25743411
-
The histone acetyltransferase p300 regulates the expression of pluripotency factors and odontogenic differentiation of human dental pulp cells.PLoS One. 2014 Jul 9;9(7):e102117. doi: 10.1371/journal.pone.0102117. eCollection 2014. PLoS One. 2014. PMID: 25007265 Free PMC article.
-
Nanog and transcriptional networks in embryonic stem cell pluripotency.Cell Res. 2007 Jan;17(1):42-9. doi: 10.1038/sj.cr.7310125. Cell Res. 2007. PMID: 17211451 Review.
-
[OCT4 and NANOG are the key genes in the system of pluripotency maintenance in mammalian cells].Genetika. 2008 Dec;44(12):1589-608. Genetika. 2008. PMID: 19178078 Review. Russian.
Cited by
-
Histone H2B monoubiquitination is a critical epigenetic switch for the regulation of autophagy.Nucleic Acids Res. 2017 Feb 17;45(3):1144-1158. doi: 10.1093/nar/gkw1025. Nucleic Acids Res. 2017. PMID: 28180298 Free PMC article.
-
Epigenetic mechanisms regulate stem cell expressed genes Pou5f1 and Gfra1 in a male germ cell line.PLoS One. 2010 Sep 14;5(9):e12727. doi: 10.1371/journal.pone.0012727. PLoS One. 2010. PMID: 20856864 Free PMC article.
-
Dual roles of histone H3 lysine 9 acetylation in human embryonic stem cell pluripotency and neural differentiation.J Biol Chem. 2015 Jan 23;290(4):2508-20. doi: 10.1074/jbc.M114.603761. Epub 2014 Dec 17. J Biol Chem. 2015. PMID: 25519907 Free PMC article.
-
The histone acetyltransferase MOF is a key regulator of the embryonic stem cell core transcriptional network.Cell Stem Cell. 2012 Aug 3;11(2):163-78. doi: 10.1016/j.stem.2012.04.023. Cell Stem Cell. 2012. PMID: 22862943 Free PMC article.
-
The stem cell--chromatin connection.Semin Cell Dev Biol. 2009 Dec;20(9):1143-8. doi: 10.1016/j.semcdb.2009.09.006. Epub 2009 Sep 16. Semin Cell Dev Biol. 2009. PMID: 19765665 Free PMC article. Review.
References
-
- Niwa, H., Miyazaki, J., and Smith, A. G. (2000) Nat. Genet. 24 372-376 - PubMed
-
- Niwa, H. (2007) Development (Camb.) 134 635-646 - PubMed
-
- Masui, S., Nakatake, Y., Toyooka, Y., Shimosato, D., Yagi, R., Takahashi, K., Okochi, H., Okuda, A., Matoba, R., Sharov, A. A., Ko, M. S., and Niwa, H. (2007) Nat. Cell Biol. 9 625-635 - PubMed
-
- Chambers, I., Colby, D., Robertson, M., Nichols, J., Lee, S., Tweedie, S., and Smith, A. (2003) Cell 113 643-655 - PubMed
-
- Mitsui, K., Tokuzawa, Y., Itoh, H., Segawa, K., Murakami, M., Takahashi, K., Maruyama, M., Maeda, M., and Yamanaka, S. (2003) Cell 113 631-642 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
