

ENDOSPERM DEFECTIVE1 Is a Novel Microtubule-Associated Protein Essential for Seed Development in Arabidopsis
- PMID: 19151224
- PMCID: PMC2648083
- DOI: 10.1105/tpc.108.061812
ENDOSPERM DEFECTIVE1 Is a Novel Microtubule-Associated Protein Essential for Seed Development in Arabidopsis
Abstract
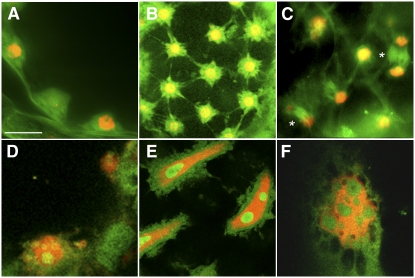
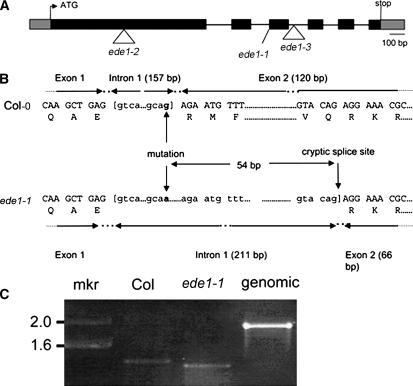
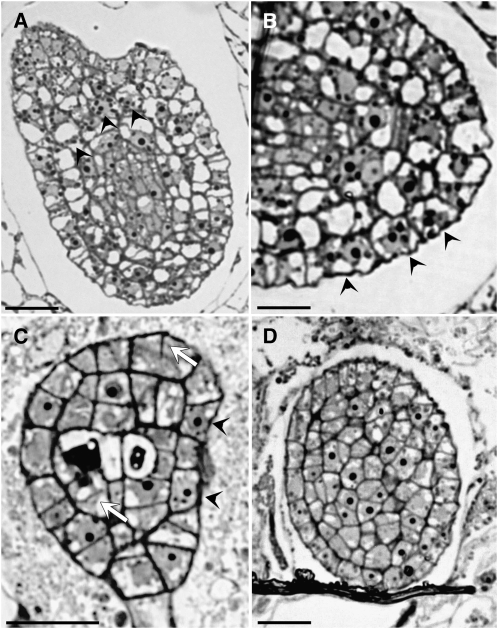
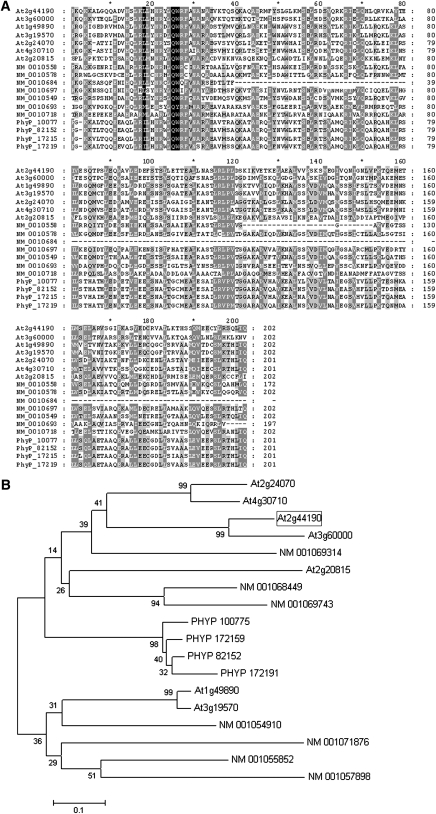
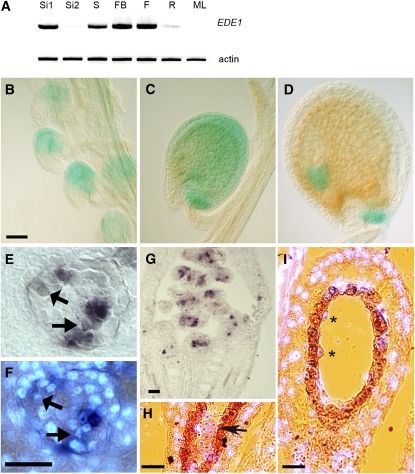
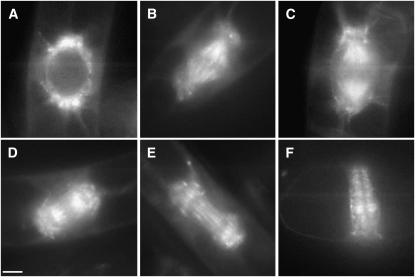
Early endosperm development involves a series of rapid nuclear divisions in the absence of cytokinesis; thus, many endosperm mutants reveal genes whose functions are essential for mitosis. This work finds that the endosperm of Arabidopsis thaliana endosperm-defective1 (ede1) mutants never cellularizes, contains a reduced number of enlarged polyploid nuclei, and features an aberrant microtubule cytoskeleton, where the specialized radial microtubule systems and cytokinetic phragmoplasts are absent. Early embryo development is substantially normal, although occasional cytokinesis defects are observed. The EDE1 gene was cloned using a map-based approach and represents the pioneer member of a conserved plant-specific family of genes of previously unknown function. EDE1 is expressed in the endosperm and embryo of developing seeds, and its expression is tightly regulated during cell cycle progression. EDE1 protein accumulates in nuclear caps in premitotic cells, colocalizes along microtubules of the spindle and phragmoplast, and binds microtubules in vitro. We conclude that EDE1 is a novel plant-specific microtubule-associated protein essential for microtubule function during the mitotic and cytokinetic stages that generate the Arabidopsis endosperm and embryo.
Figures


References
-
- Alonso, J.M., et al. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301 653–657. - PubMed
-
- Altschul, S.F., Gish, W., Miller, W., Myers, E.W., and Lipman, D.J. (1990). Basic local alignment search tool. J. Mol. Biol. 215 403–410. - PubMed
-
- Assaad, F.F., Mayer, U., Wanner, G., and Jürgens, G. (1996). The KEULE gene is involved in cytokinesis in Arabidopsis. Mol. Gen. Genet. 253 267–277. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
