

Cytoplasmic penetration and persistent infection of mammalian cells by polyglutamine aggregates
- PMID: 19151706
- PMCID: PMC2757079
- DOI: 10.1038/ncb1830
Cytoplasmic penetration and persistent infection of mammalian cells by polyglutamine aggregates
Abstract
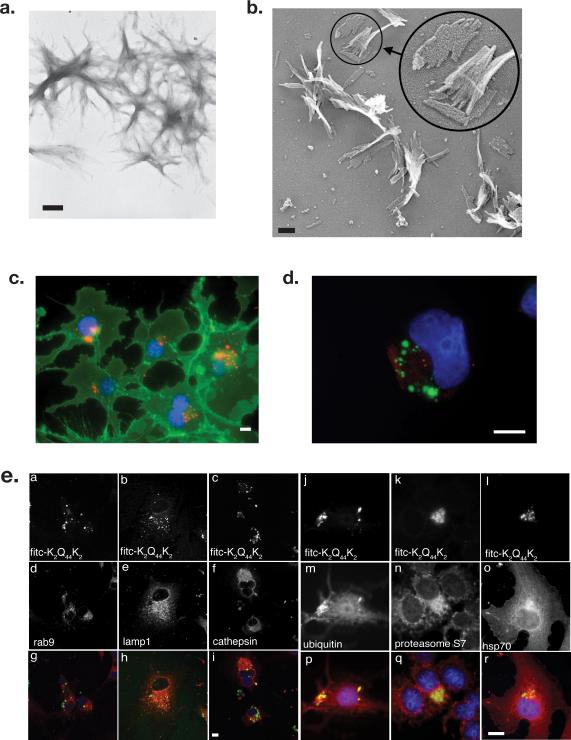
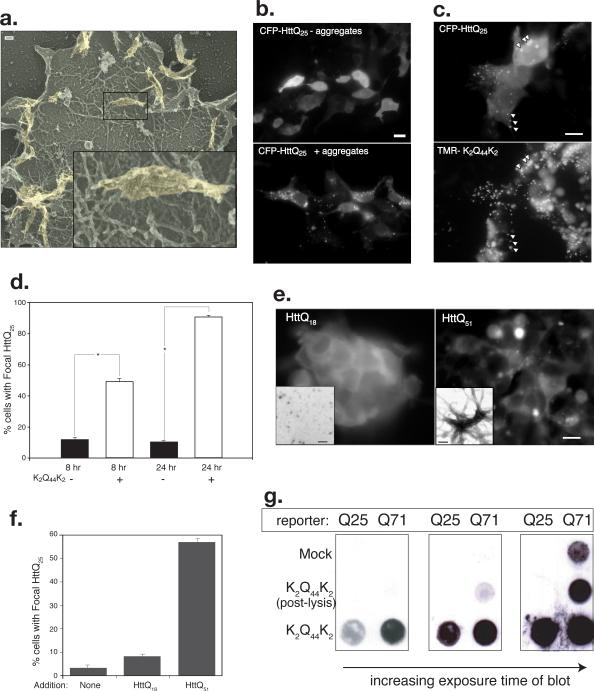
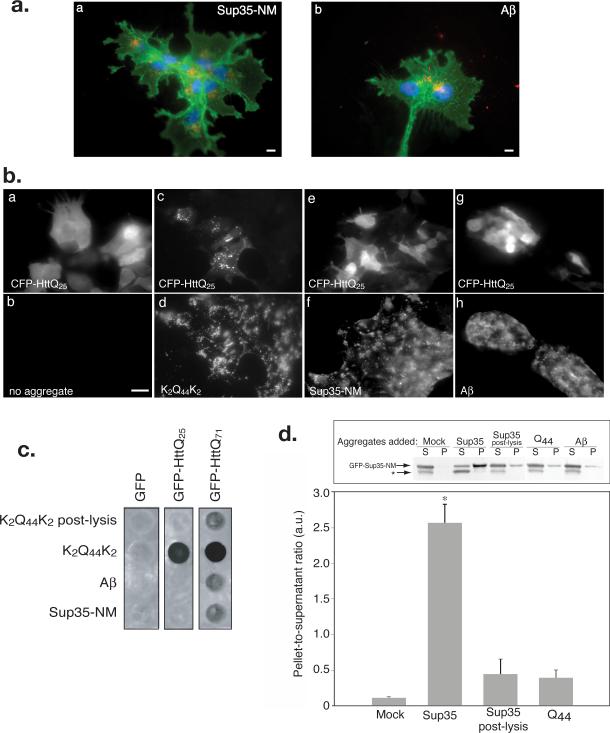
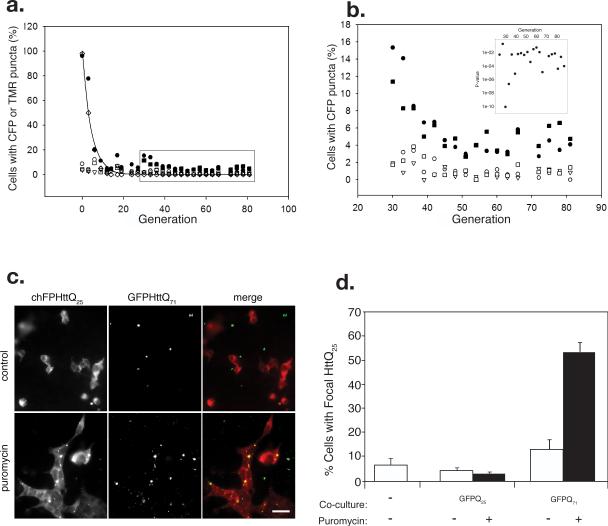
Sequence-specific nucleated protein aggregation is closely linked to the pathogenesis of most neurodegenerative diseases and constitutes the molecular basis of prion formation. Here we report that fibrillar polyglutamine peptide aggregates can be internalized by mammalian cells in culture where they gain access to the cytosolic compartment and become co-sequestered in aggresomes together with components of the ubiquitin-proteasome system and cytoplasmic chaperones. Remarkably, these internalized fibrillar aggregates are able to selectively recruit soluble cytoplasmic proteins with which they share homologous but not heterologous amyloidogenic sequences, and to confer a heritable phenotype on cells expressing the homologous amyloidogenic protein from a chromosomal locus.
Figures
References
-
- Chiti F, Dobson CM. Protein misfolding, functional amyloid, and human disease. Annu Rev Biochem. 2006;75:333–366. - PubMed
-
- Taylor JP, Hardy J, Fischbeck KH. Toxic proteins in neurodegenerative disease. Science. 2002;296:1991–1995. - PubMed
-
- Carrell RW, Lomas DA. Conformational disease. Lancet. 1997;350:134–138. - PubMed
-
- Pepys MB. Amyloidosis. Annu Rev Med. 2006;57:223–241. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
