

A distinct class of small RNAs arises from pre-miRNA-proximal regions in a simple chordate
- PMID: 19151725
- PMCID: PMC2746024
- DOI: 10.1038/nsmb.1536
A distinct class of small RNAs arises from pre-miRNA-proximal regions in a simple chordate
Abstract
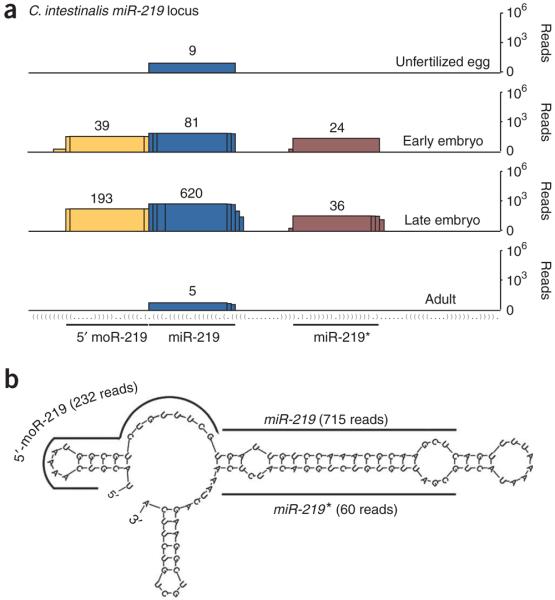
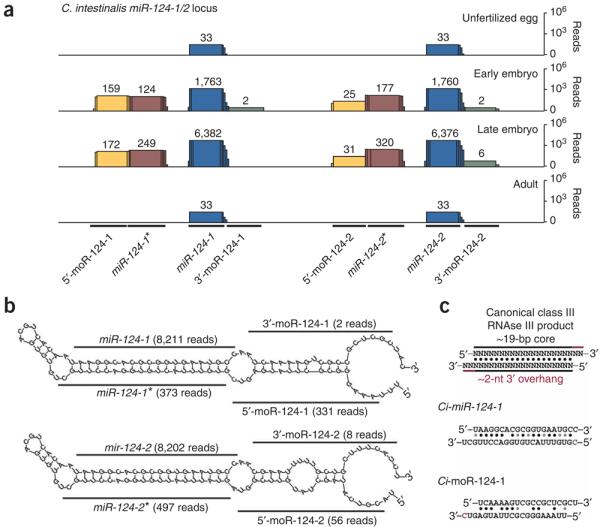
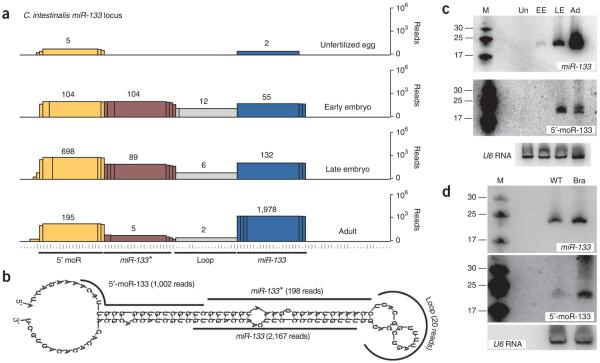
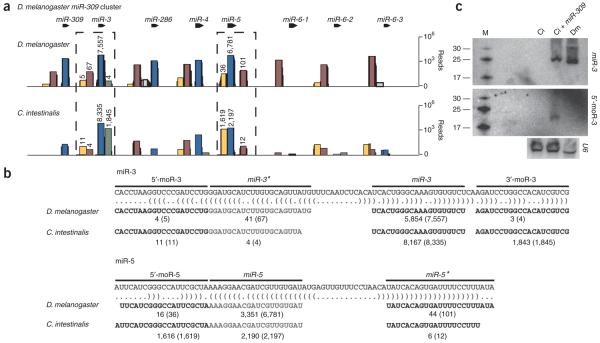
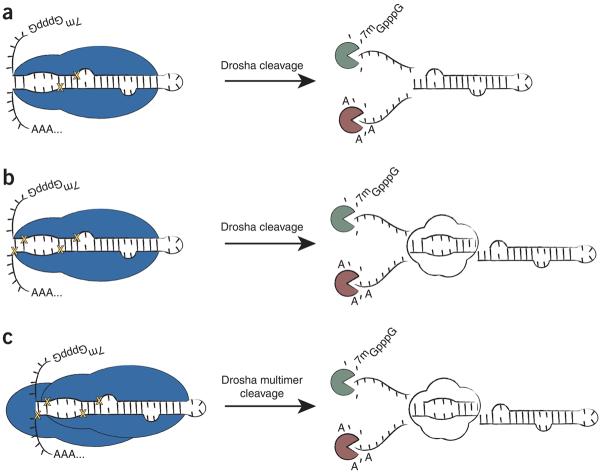
MicroRNAs (miRNAs) have been implicated in various cellular processes. They are thought to function primarily as inhibitors of gene activity by attenuating translation or promoting mRNA degradation. A typical miRNA gene produces a predominant approximately 21-nucleotide (nt) RNA (the miRNA) along with a less abundant miRNA(*) product. We sought to identify miRNAs from the simple chordate Ciona intestinalis through comprehensive sequencing of small RNA libraries created from different developmental stages. Unexpectedly, half of the identified miRNA loci encode up to four distinct, stable small RNAs. The additional RNAs, miRNA-offset RNAs (moRs), are generated from sequences immediately adjacent to the predicted approximately 60-nt pre-miRNA. moRs seem to be produced by RNAse III-like processing, are approximately 20 nt long and, like miRNAs, are observed at specific developmental stages. We present evidence suggesting that the biogenesis of moRs results from an intrinsic property of the miRNA processing machinery in C. intestinalis.
Figures
References
-
- Ambros V. The functions of animal microRNAs. Nature. 2004;431:350–355. - PubMed
-
- Zamore PD, Haley B. Ribo-gnome: the big world of small RNAs. Science. 2005;309:1519–1524. - PubMed
-
- Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116:281–297. - PubMed
-
- Lau NC, Lim LP, Weinstein EG, Bartel DP. An abundant class of tiny RNAs with probable regulatory roles in Caenorhabditis elegans. Science. 2001;294:858–862. - PubMed
-
- Pasquinelli AE, et al. Conservation of the sequence and temporal expression of let-7 heterochronic regulatory RNA. Nature. 2000;408:86–89. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
