

TRPP2 channels regulate apoptosis through the Ca2+ concentration in the endoplasmic reticulum
- PMID: 19153608
- PMCID: PMC2657577
- DOI: 10.1038/emboj.2008.307
TRPP2 channels regulate apoptosis through the Ca2+ concentration in the endoplasmic reticulum
Abstract
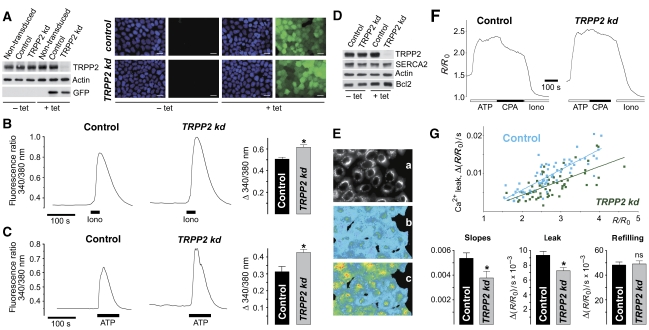
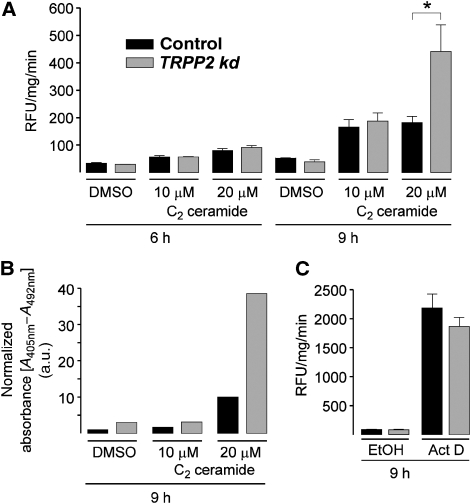
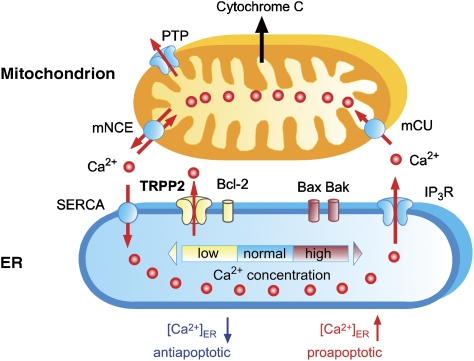
Ca(2+) is an important signalling molecule that regulates multiple cellular processes, including apoptosis. Although Ca(2+) influx through transient receptor potential (TRP) channels in the plasma membrane is known to trigger cell death, the function of intracellular TRP proteins in the regulation of Ca(2+)-dependent signalling pathways and apoptosis has remained elusive. Here, we show that TRPP2, the ion channel mutated in autosomal dominant polycystic kidney disease (ADPKD), protects cells from apoptosis by lowering the Ca(2+) concentration in the endoplasmic reticulum (ER). ER-resident TRPP2 counteracts the activity of the sarcoendoplasmic Ca(2+) ATPase by increasing the ER Ca(2+) permeability. This results in diminished cytosolic and mitochondrial Ca(2+) signals upon stimulation of inositol 1,4,5-trisphosphate receptors and reduces Ca(2+) release from the ER in response to apoptotic stimuli. Conversely, knockdown of TRPP2 in renal epithelial cells increases ER Ca(2+) release and augments sensitivity to apoptosis. Our findings indicate an important function of ER-resident TRPP2 in the modulation of intracellular Ca(2+) signalling, and provide a molecular mechanism for the increased apoptosis rates in ADPKD upon loss of TRPP2 channel function.
Figures
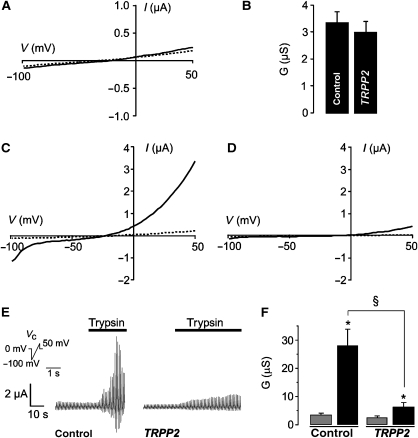
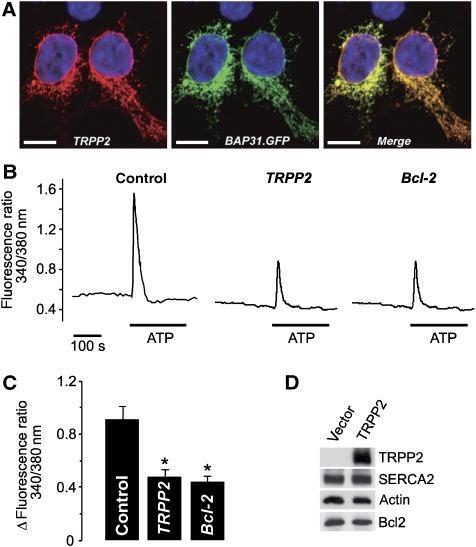
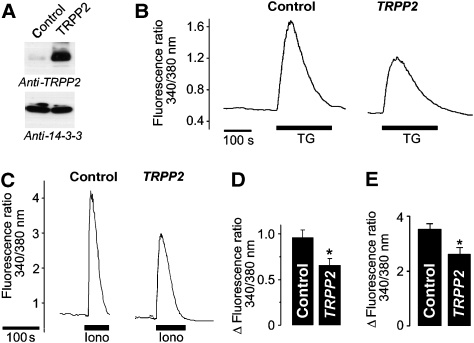
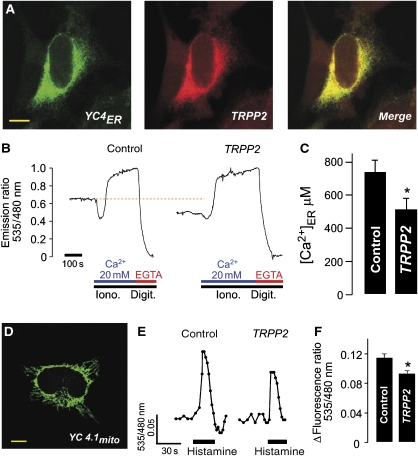
References
-
- Arnaudeau S, Frieden M, Nakamura K, Castelbou C, Michalak M, Demaurex N (2002) Calreticulin differentially modulates calcium uptake and release in the endoplasmic reticulum and mitochondria. J Biol Chem 277: 46696–46705 - PubMed
-
- Arnaudeau S, Kelley WL, Walsh JV Jr, Demaurex N (2001) Mitochondria recycle Ca2+ to the endoplasmic reticulum and prevent the depletion of neighboring endoplasmic reticulum regions. J Biol Chem 276: 29430–29439 - PubMed
-
- Berridge MJ, Lipp P, Bootman MD (2000) The versatility and universality of calcium signalling. Nat Rev Mol Cell Biol 1: 11–21 - PubMed
-
- Boletta A, Germino GG (2003) Role of polycystins in renal tubulogenesis. Trends Cell Biol 13: 484–492 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
