

Control of translation initiation involves a factor-induced rearrangement of helix 44 of 16S ribosomal RNA
- PMID: 19154330
- PMCID: PMC3647337
- DOI: 10.1111/j.1365-2958.2009.06598.x
Control of translation initiation involves a factor-induced rearrangement of helix 44 of 16S ribosomal RNA
Abstract
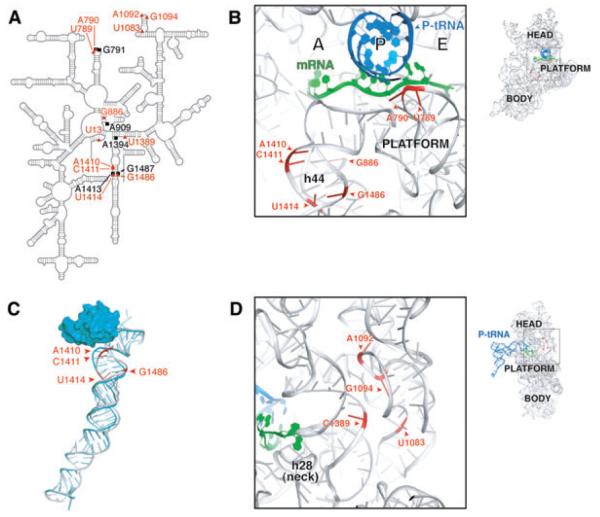
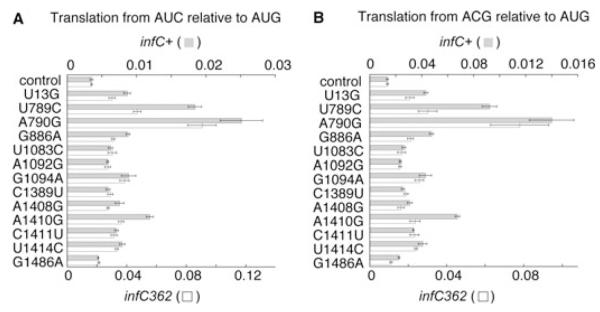
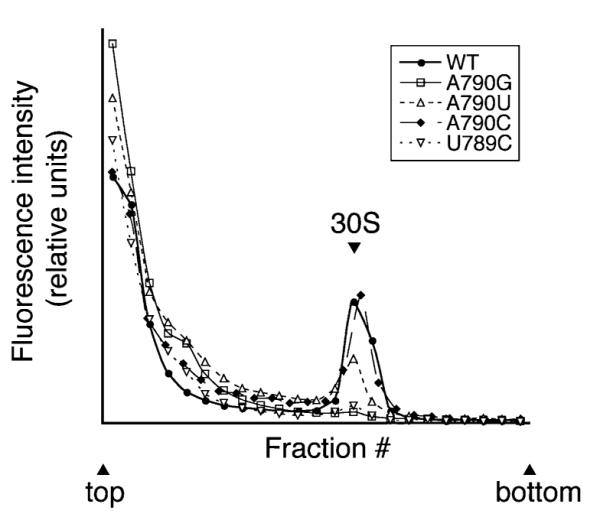
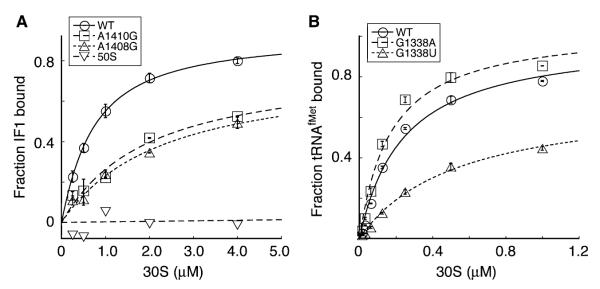
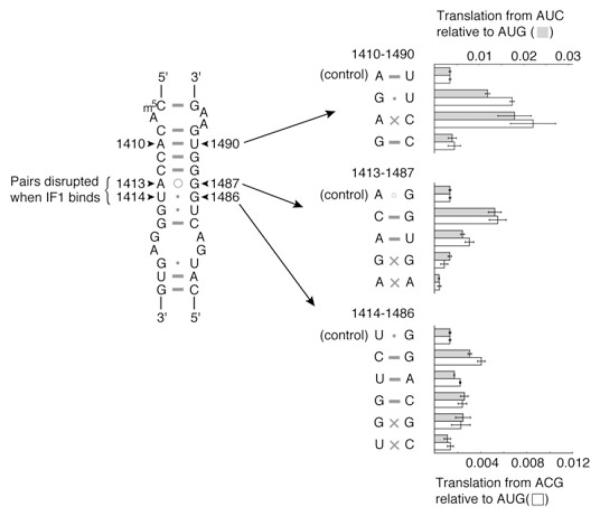
Initiation of translation involves recognition of the start codon by the initiator tRNA in the 30S subunit. To investigate the role of ribosomal RNA (rRNA) in this process, we isolated a number of 16S rRNA mutations that increase translation from the non-canonical start codon AUC. These mutations cluster to distinct regions that overlap remarkably well with previously identified class III protection sites and implicate both IF1 and IF3 in start codon selection. Two mutations map to the 790 loop and presumably act by inhibiting IF3 binding. Another cluster of mutations surrounds the conserved A1413(o)G1487 base pair of helix 44 in a region known to be distorted by IF1 and IF3. Site-directed mutagenesis in this region confirmed that this factor-induced rearrangement of helix 44 helps regulate initiation fidelity. A third cluster of mutations maps to the neck of the 30S subunit, suggesting that the dynamics of the head domain influences translation initiation. In addition to identifying mutations that decrease fidelity, we found that many P-site mutations increase the stringency of start codon selection. These data provide evidence that the interaction between the initiator tRNA and the 30S P site is tuned to balance efficiency and accuracy during initiation.
Figures
Similar articles
-
Characterization of 16S rRNA mutations that decrease the fidelity of translation initiation.RNA. 2007 Dec;13(12):2348-55. doi: 10.1261/rna.715307. Epub 2007 Oct 17. RNA. 2007. PMID: 17942743 Free PMC article.
-
Initiation factor 3-induced structural changes in the 30 S ribosomal subunit and in complexes containing tRNA(f)(Met) and mRNA.J Mol Biol. 2000 Jun 9;299(3):615-28. doi: 10.1006/jmbi.2000.3774. J Mol Biol. 2000. PMID: 10835272
-
Structural analysis of noncanonical translation initiation complexes.J Biol Chem. 2024 Oct;300(10):107743. doi: 10.1016/j.jbc.2024.107743. Epub 2024 Sep 1. J Biol Chem. 2024. PMID: 39222680 Free PMC article.
-
Role of the initiation factor 3 in the fidelity of initiator tRNA selection on ribosome.IUBMB Life. 2025 Jan;77(1):e2927. doi: 10.1002/iub.2927. Epub 2024 Nov 23. IUBMB Life. 2025. PMID: 39578968 Review.
-
Eukaryotic initiator tRNA: finely tuned and ready for action.FEBS Lett. 2010 Jan 21;584(2):396-404. doi: 10.1016/j.febslet.2009.11.047. FEBS Lett. 2010. PMID: 19925799 Free PMC article. Review.
Cited by
-
Real-time assembly landscape of bacterial 30S translation initiation complex.Nat Struct Mol Biol. 2012 May 6;19(6):609-15. doi: 10.1038/nsmb.2285. Nat Struct Mol Biol. 2012. PMID: 22562136
-
Large-Scale Movements of IF3 and tRNA during Bacterial Translation Initiation.Cell. 2016 Sep 22;167(1):133-144.e13. doi: 10.1016/j.cell.2016.08.074. Cell. 2016. PMID: 27662086 Free PMC article.
-
Role of helix 44 of 16S rRNA in the fidelity of translation initiation.RNA. 2012 Mar;18(3):485-95. doi: 10.1261/rna.031203.111. Epub 2012 Jan 25. RNA. 2012. PMID: 22279149 Free PMC article.
-
Conformational Response of 30S-bound IF3 to A-Site Binders Streptomycin and Kanamycin.Antibiotics (Basel). 2016 Dec 13;5(4):38. doi: 10.3390/antibiotics5040038. Antibiotics (Basel). 2016. PMID: 27983590 Free PMC article.
-
Directional transition from initiation to elongation in bacterial translation.Nucleic Acids Res. 2015 Dec 15;43(22):10700-12. doi: 10.1093/nar/gkv869. Epub 2015 Sep 3. Nucleic Acids Res. 2015. PMID: 26338773 Free PMC article.
References
-
- Antoun A, Pavlov MY, Lovmar M, Ehrenberg M. How initiation factors maximize the accuracy of tRNA selection in initiation of bacterial protein synthesis. Mol Cell. 2006a;23:183–193. - PubMed
-
- Boelens R, Gualerzi CO. Structure and function of bacterial initiation factors. Curr Protein Pept Sci. 2002;3:107–119. - PubMed
-
- Brombach M, Pon CL. The unusual translational initiation codon AUU limits the expression of the infC (initiation factor IF3) gene of Escherichia coli. Mol Gen Genet. 1987;208:94–100. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
