

Potent anti-inflammatory effects of two quinolinedione compounds, OQ1 and OQ21, mediated by dual inhibition of inducible NO synthase and cyclooxygenase-2
- PMID: 19154436
- PMCID: PMC2697840
- DOI: 10.1111/j.1476-5381.2008.00028.x
Potent anti-inflammatory effects of two quinolinedione compounds, OQ1 and OQ21, mediated by dual inhibition of inducible NO synthase and cyclooxygenase-2
Abstract
Background and purpose: Inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) have been suggested as key components in various inflammatory diseases. Here we examined the effects of new quinolinedione derivatives, 6-(4-fluorophenyl)-amino-5,8-quinolinedione (OQ1) and 6-(2,3,4-trifluorophenyl)-amino-5,8-quinolinedione (OQ21) on activity and expression of iNOS and COX-2 to explore their anti-inflammatory properties.
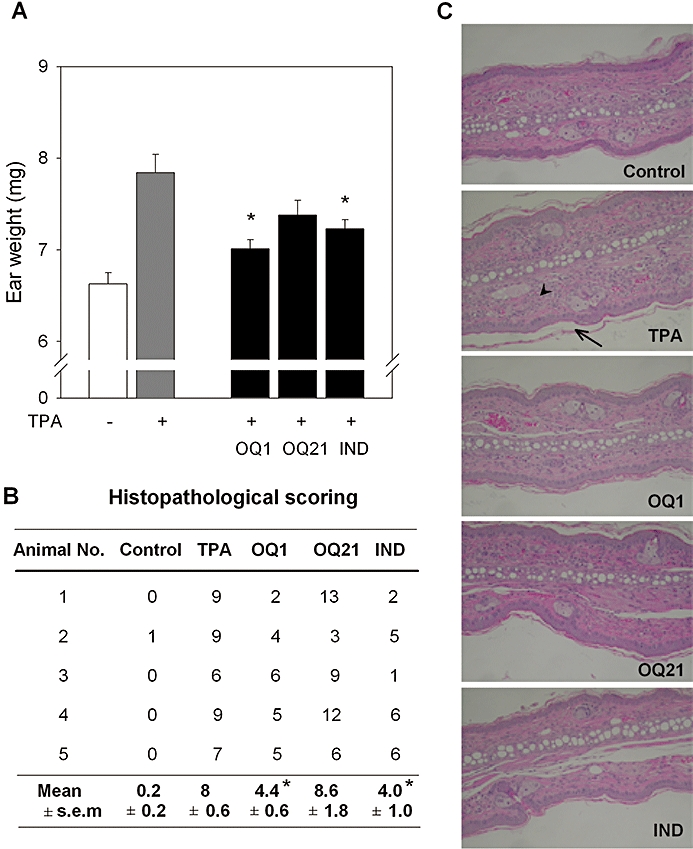
Experimental approach: The effects of OQ1 and OQ21 were assessed on lipopolysaccharide (LPS)-induced iNOS and COX-2 in murine macrophage cell line (RAW264.7), along with isolated enzyme assays to measure enzyme inhibition. Nuclear factor-kappaB (NFkappaB) activation pathways were investigated to elucidate mechanisms underlying OQ-mediated suppression of the expression of iNOS and COX-2. In vivo anti-inflammatory activities of OQ compounds were evaluated in mouse ear oedema, induced by topical 12-O-tetradecanoylphorbol-13-acetate (TPA).
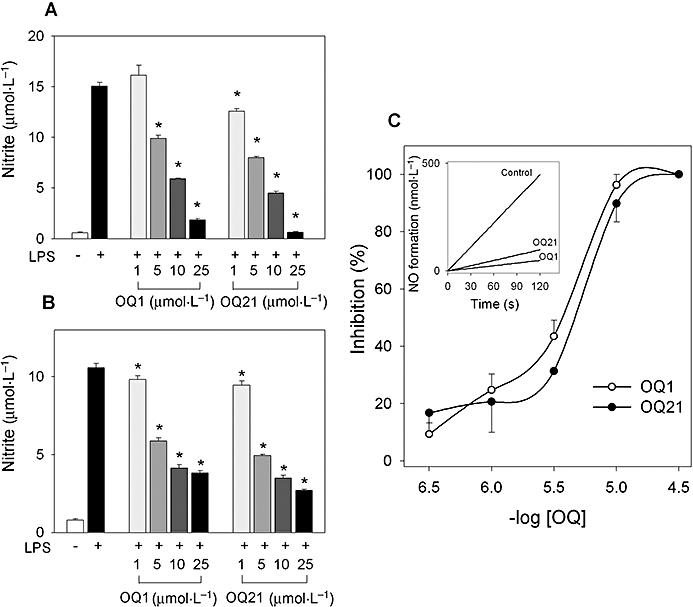
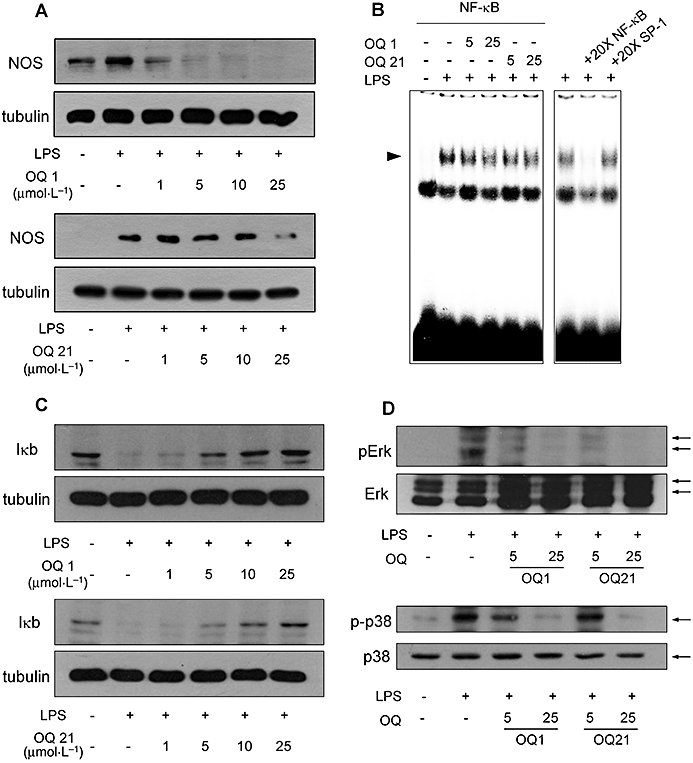
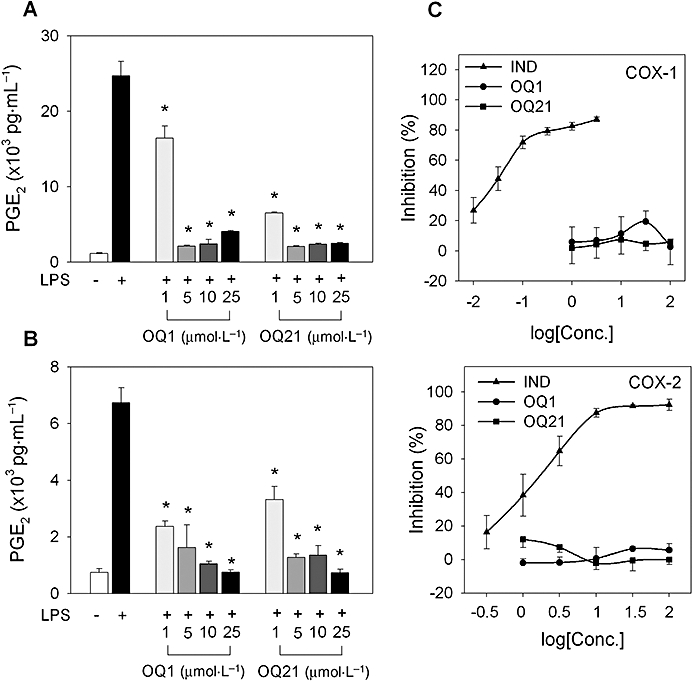
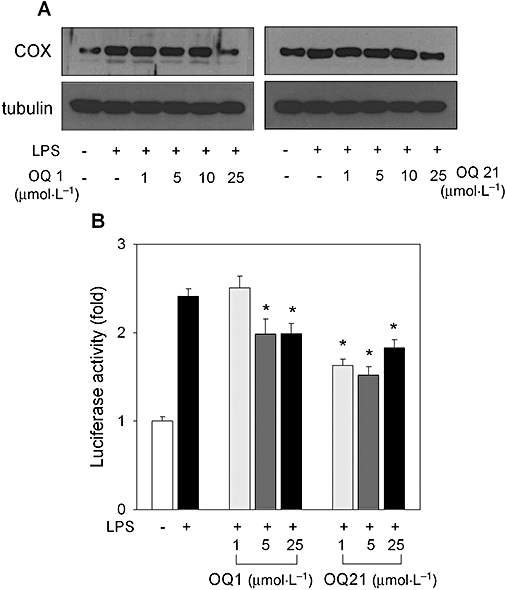
Key results: LPS-induced NO production in RAW264.7 cells was inhibited by OQ1 and OQ21 through the attenuation of iNOS expression as well as iNOS activity. Down-regulation of iNOS followed blocking of NFkappaB activation, as assessed by inhibitory kappaB degradation and electrophoretic mobility shift assay for NFkappaB. Synthesis and accumulation of prostaglandin E(2) were also suppressed by OQ1 and OQ21. LPS-induced COX-2 expression and cellular COX-2 activities were attenuated by OQ1 and OQ21. Consistent with these results, OQ1 showed potent anti-inflammatory effects in mouse ear oedema induced by TPA.
Conclusions and implications: The novel quinolinedione derivatives, OQ1 and OQ21, showed potent anti-inflammatory activity through dual inhibitory effects on iNOS and COX-2, suggesting that OQ derivatives might provide a new therapeutic modality for chronic inflammatory diseases, refractory to conventional drug therapies.
Figures
Similar articles
-
Aciculatin inhibits lipopolysaccharide-mediated inducible nitric oxide synthase and cyclooxygenase-2 expression via suppressing NF-κB and JNK/p38 MAPK activation pathways.J Biomed Sci. 2011 May 6;18(1):28. doi: 10.1186/1423-0127-18-28. J Biomed Sci. 2011. PMID: 21548916 Free PMC article.
-
Anti-inflammatory activity of 4-methoxyhonokiol is a function of the inhibition of iNOS and COX-2 expression in RAW 264.7 macrophages via NF-kappaB, JNK and p38 MAPK inactivation.Eur J Pharmacol. 2008 May 31;586(1-3):340-9. doi: 10.1016/j.ejphar.2008.02.044. Epub 2008 Feb 26. Eur J Pharmacol. 2008. PMID: 18378223
-
Anti-inflammatory effect of allylpyrocatechol in LPS-induced macrophages is mediated by suppression of iNOS and COX-2 via the NF-kappaB pathway.Int Immunopharmacol. 2008 Sep;8(9):1264-71. doi: 10.1016/j.intimp.2008.05.003. Epub 2008 Jun 6. Int Immunopharmacol. 2008. PMID: 18602073
-
Inhibition of inducible NO synthase, cyclooxygenase-2 and interleukin-1beta by torilin is mediated by mitogen-activated protein kinases in microglial BV2 cells.Br J Pharmacol. 2009 Mar;156(6):933-40. doi: 10.1111/j.1476-5381.2009.00022.x. Br J Pharmacol. 2009. PMID: 19298258 Free PMC article.
-
Synthesis, Biological Evaluation, and Molecular Docking Study of 3-Amino and 3-Hydroxy-seco A Derivatives of α-Amyrin and 3-Epilupeol as Inhibitors of COX-2 Activity and NF-kB Activation.J Nat Prod. 2022 Apr 22;85(4):787-803. doi: 10.1021/acs.jnatprod.1c00827. Epub 2022 Feb 17. J Nat Prod. 2022. PMID: 35175765 Review.
Cited by
-
Exploring the Therapeutic Marvels: A Comprehensive Review on the Biological Potential of Quinoline-5,8-Dione.Med Chem. 2024;20(4):385-396. doi: 10.2174/0115734064287677231215070816. Med Chem. 2024. PMID: 38173200 Review.
-
Anti-inflammatory and arthritic effects of thiacremonone, a novel sulfur compound isolated from garlic via inhibition of NF-kappaB.Arthritis Res Ther. 2009;11(5):R145. doi: 10.1186/ar2819. Epub 2009 Sep 30. Arthritis Res Ther. 2009. PMID: 19788760 Free PMC article.
-
5,8-Quinolinedione Scaffold as a Promising Moiety of Bioactive Agents.Molecules. 2019 Nov 14;24(22):4115. doi: 10.3390/molecules24224115. Molecules. 2019. PMID: 31739496 Free PMC article. Review.
-
The effect of some 4,2 and 5,2 bisthiazole derivatives on nitro-oxidative stress and phagocytosis in acute experimental inflammation.Molecules. 2014 Jul 2;19(7):9240-56. doi: 10.3390/molecules19079240. Molecules. 2014. PMID: 24991757 Free PMC article.
-
A one-pot three-component reaction involving 2-aminochromone in aqueous micellar medium: a green synthesis of hexahydrochromeno[2,3-b]quinolinedione.Mol Divers. 2015 Aug;19(3):541-9. doi: 10.1007/s11030-015-9573-7. Epub 2015 Mar 11. Mol Divers. 2015. PMID: 25758539
References
-
- Ajizian SJ, English BK, Meals EA. Specific inhibitors of p38 and extracellular signal-regulated kinase mitogen-activated protein kinase pathways block inducible nitric oxide synthase and tumor necrosis factor accumulation in murine macrophages stimulated with lipopolysaccharide and interferon-gamma. J Infect Dis. 1999;179:939–944. - PubMed
-
- Chen CH, Sheu MT, Chen TF, Wang YC, Hou WC, Liu DZ, et al. Suppression of endotoxin-induced proinflammatory responses by citrus pectin through blocking LPS signaling pathways. Biochem Pharmacol. 2006;72:1001–1009. - PubMed
-
- Chen YC, Shen SC, Lee WR, Hou WC, Yang LL, Lee TJ. Inhibition of nitric oxide synthase inhibitors and lipopolysaccharide induced inducible NOS and cyclooxygenase-2 gene expressions by rutin, quercetin, and quercetin pentaacetate in RAW 264.7 macrophages. J Cell Biochem. 2001;82:537–548. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
