

Unattached kinetochores catalyze production of an anaphase inhibitor that requires a Mad2 template to prime Cdc20 for BubR1 binding
- PMID: 19154722
- PMCID: PMC2655205
- DOI: 10.1016/j.devcel.2008.11.005
Unattached kinetochores catalyze production of an anaphase inhibitor that requires a Mad2 template to prime Cdc20 for BubR1 binding
Abstract
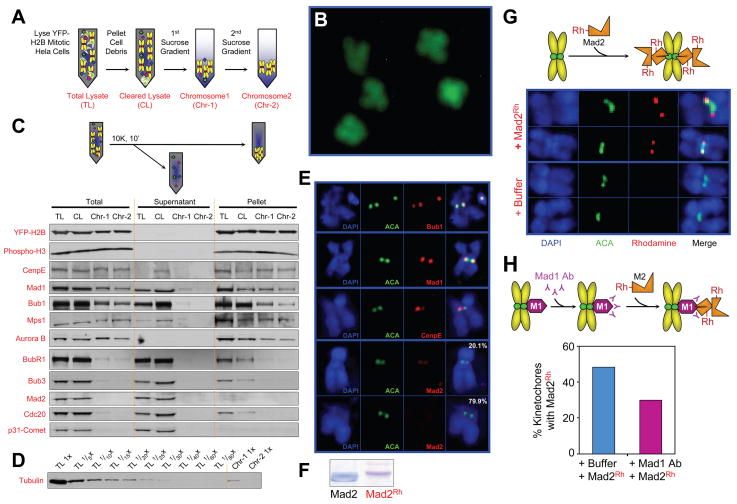
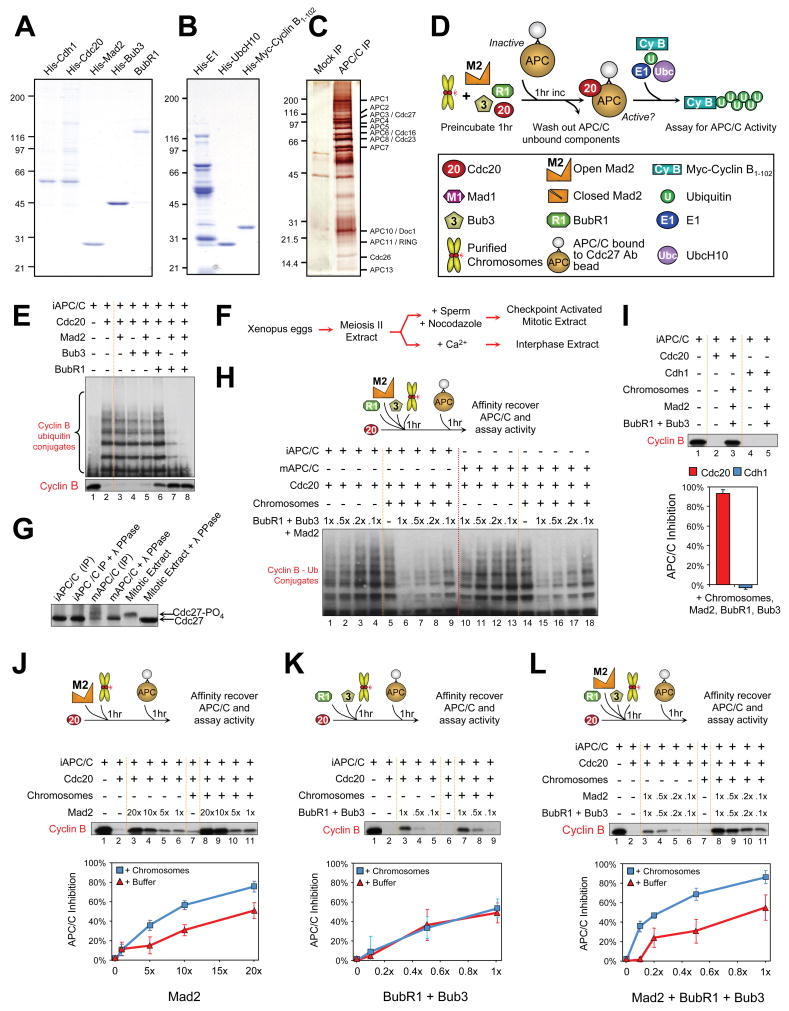
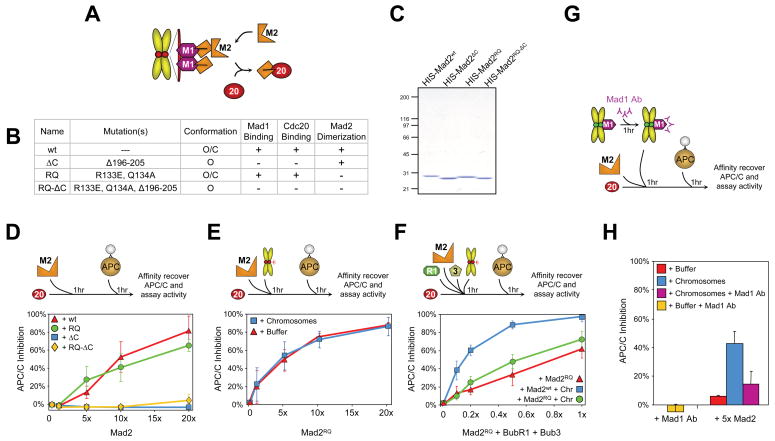
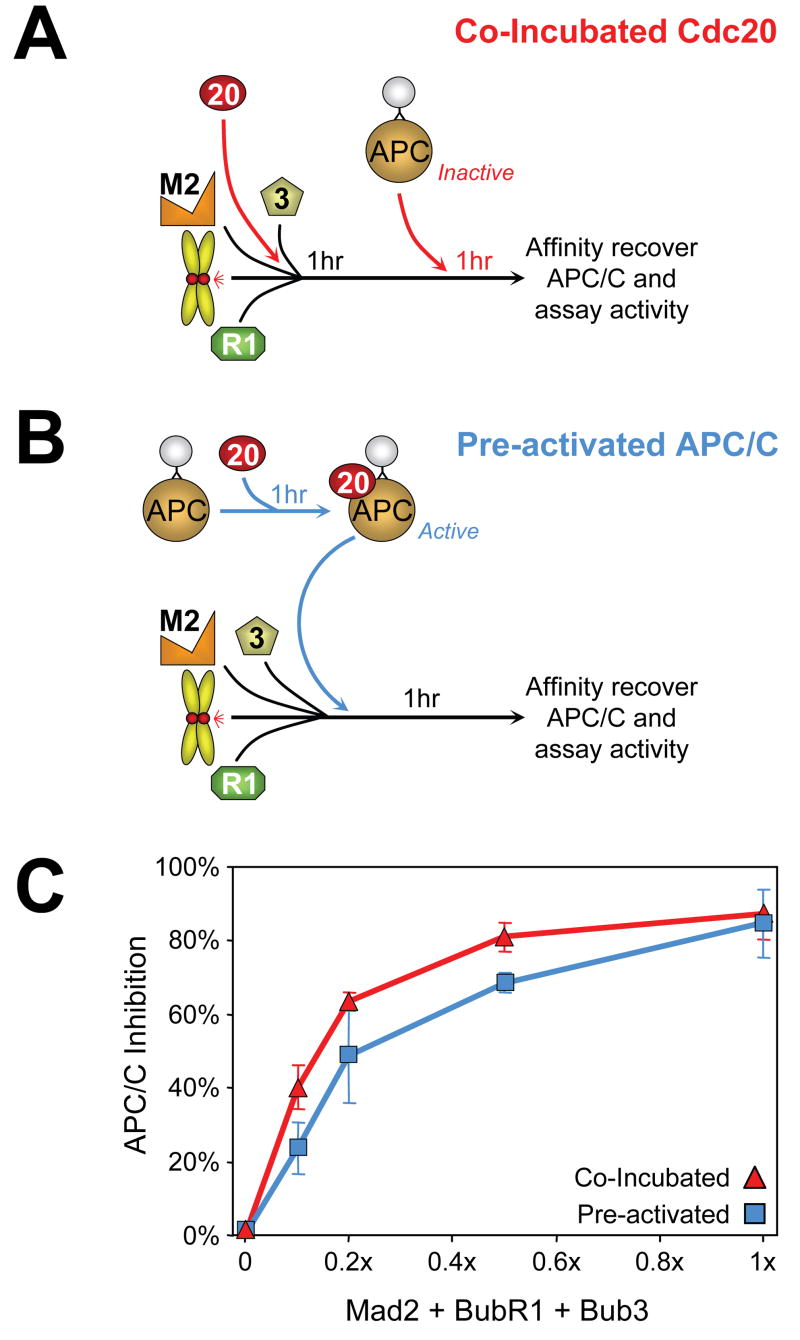
Premature anaphase onset is prevented by the mitotic checkpoint through production of a "wait anaphase" inhibitor(s) that blocks recognition of cyclin B and securin by Cdc20-activated APC/C, an E3 ubiquitin ligase that targets them for destruction. Using physiologically relevant levels of Mad2, Bub3, BubR1, and Cdc20, we demonstrate that unattached kinetochores on purified chromosomes catalytically generate a diffusible Cdc20 inhibitor or inhibit Cdc20 already bound to APC/C. Furthermore, the chromosome-produced inhibitor requires both recruitment of Mad2 by Mad1 that is stably bound at unattached kinetochores and dimerization-competent Mad2. We show that purified chromosomes promote BubR1 binding to APC/C-Cdc20 by acting directly on Mad2, but not BubR1. Our results support a model in which immobilized Mad1/Mad2 at kinetochores provides a template for initial assembly of Mad2 bound to Cdc20 that is then converted to a final mitotic checkpoint inhibitor with Cdc20 bound to BubR1.
Figures
Comment in
-
Relaying the checkpoint signal from kinetochore to APC/C.Dev Cell. 2009 Jan;16(1):6-8. doi: 10.1016/j.devcel.2008.12.008. Dev Cell. 2009. PMID: 19154713
References
-
- Acquaviva C, Herzog F, Kraft C, Pines J. The anaphase promoting complex/cyclosome is recruited to centromeres by the spindle assembly checkpoint. Nat Cell Biol. 2004;6:892–898. - PubMed
-
- Brown KD, Wood KW, Cleveland DW. The kinesin-like protein CENP-E is kinetochore-associated throughout poleward chromosome segregation during anaphase-A. J Cell Sci. 1996;109(Pt 5):961–969. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
